Besin ağı
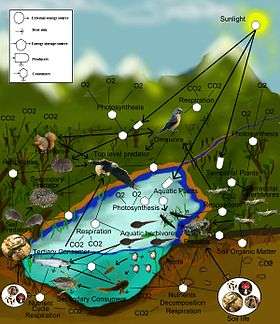
Besin ağı ya da besin döngüsü besin zincirlerinin doğal olarak bağlaşmasıdır ve genellikle ekolojik bir topluluk içinde neyin ne ile beslendiğini gösteren grafiksel bir gösterimdir. Çevrebilimciler tüm yaşam biçimleri kabaca trofik düzey adı verilen iki kategoride sınıflandırırlar: ototroflar ve heterotroflar. Ototroflar büyümek, gelişmek ve üremek için mineraller ve karbon dioksit gibi gazlardan oluşan inorganik maddelerden organik madde üreterek kendi besinlerini sağlarlar. Bu kimyasal tepkimelerin gerektirdiği enerji güneşten çoğunlukla fotosentez yoluyla elde edilir. Hidrotermal bacalar ve kaplıcalar az da olsa güneşin yanında diğer enerji kaynaklarıdır. Trofik düzeyler karbon gereksinimlerini yalnızca atmosferden elde eden tam ototroflardan, organik maddeyi atmosferden elde etmenin yanısıra diğer kaynakları da kullanan etçil bitkiler gibi miksotroflara ve organik madde elde etmek için beslenmek zorunda olan tam heterotroflara kadar uzanır. Besin ağında besin zincirleri heterotrofların hangi ototroflar ya da heterotroflar ile beslendiğini gösteren bağlantılar ile gösterilir. Besin ağı bir ekosistemi değiş-tokuş yapan birleşik bir sistem olarak çeşitli beslenme yöntemlerinin basit olarak tasvir edilmesidir. Kabaca otçul, etçil, leşçil ve parazitik olarak ayrılabilen değişik beslenme ilişkileri vardır. Heterotroflar tarafından yenilen şekerler gibi bazı organik maddeler enerji sağlar. Siyanobakterilerden sekoyaya ve virüslerden mavi balinaya kadar ototroflar ve heterotroflar mikroskobik boyuttan tonlarca ağırlığa kadar her boyutta bulunmaktadırlar.
Charles Elton besin döngüsü, besin zinciri ve besin boyutu kavramlarına 1927 yılında yayımlanan "Animal Ecology" (Hayvan Ekolojisi) adlı kitabıyla öncülük etmiştir; daha sonra gelen ekoloji ile ilgili metinlerde "besin döngüsü" terimi "besin ağı" ile değiştirilmiştir. Elton'ın türleri sınıflandırdığı işlevsel gruplar Raymond Lindeman'ın 1942 yılında trofik dinamikleri üzerine yayımladığı önemli makalesine temel olmuştur. Lindeman bu makalesinde trofik sınıflandırma sistemi içinde ayrıştırıcı organizmaların önemini vurgulamıştır. Besin ağı terimi tarihi olarak Charles Darwin'in eserlerinde geçen "girift yamaç", "yaşam ağı" ve "karmaşık ilişkiler ağı" gibi terimlere dayanmaktadır. Hatta daha önceden John Bruckner 1768 yılında doğayı "süregelen bir yaşam ağı" olarak tanımlamıştır.
Besin ağları, aynı avlara sahip olan ve aynı avcılar tarafından avlanan çok sayıda türü işlevsel grup olarak trofik türlere indirgeyerek gerçek ekosistemlerin sınırlı olarak tasvirleridir. Çevrebilimciler bu sadeleştirmeleri trofik ya da üretici-tüketici sistem dinamiklerinin kantitatif modellemesinde kullanırlar. Bu modellemeler sayesinde gerçek besin ağı örgülerinin yapısında yer alan genel örüntüleri ölçebilmekte ve değerlendirebilmektedirler. Bu çalışmalar sonucunda besn ağlarının topolojisinde tesadüfî olmayan nitelikleri tanımlamışlardır. Meta analizde kullanılan yayımlanmış besin ağı örnekleri içerisinde eksikler bulunan çeşitli kalite düzeyindedir. Ancak topluluk aüları üzerine yapılan ampirik çalışmalar artmaktadır ve ağ teorisi kullanılarak besin ağlarının matematiksel analizi tüm besin ağlarında görülen ortak özelikleri ortaya çıkarmıştır. Örneğin üstel kanun besin ağı topolojisi içinde avcı-av bağlantılarını ve tür zenginliğini öngörmek için kullanılmaktadır.
Besin ağı taksonomisi
Besin ağı haritasındaki bağlantılar ekolojik bir toplulukta neyin ne ile beslendiğini gösteren ilişkilerdir. Çevrebilimciler tüm canlıları kabaca ototrof ve heterotrof olarak iki trofik düzeye ayırırlar. Ototroflar ya güneş enerjisini kullanmadan kimyasal olarak ya da güneş enerjisini kullanarak fotosentez yoluyla ürettikleri biyokütle enerjisini metabolik solunum sırasında kullanırlar. Heterotroflar ise üretmekten çok biyokütle enerjisini tüketerek metabolize eder ve büyüyerek ikincil üretim düzeylerine katkıda bulunurlar. Bir besin ağı kendi besinlerini üreten temel üretici ototroflardan aldıkları besinlerle beslenen polifajik heterotrof tüketiciler topluluğu arasında bulunan enerji akışı döngüsünü tasvir eder.[1][2][3]
Bir besin ağında yer alan temel türler avı olmayan ototroflar ve toprakta, biyofilmde ya da perifitonda bulunan ayrıştırıcılardan oluşan saprofitik detritivorlardan oluşabilir. Ağ içindeki beslenme bağlarına trofik bağ adı verilir. Tüketici başına trofik bağ sayısı besin ağının karmaşıklığının bir ölçüsüdür. Besin zincirleri besin ağının trofik bağlarında iç içe geçmiş olarak bulunur. Besin zincirleri temel üreticiden genelde büyük bir etçil olan süper avcıya kadar tekdüze tüketicileri izleyen doğrusal beslenme bağlarıdır.[4][5][6]
Besin ağındaki bağlar trofik tür adı verilen biyolojik takson topluluklarından oluşan düğüm noktalarına bağlanır. Trofik türler besin ağı içinde aynı av ve avcılara sahip olan türlerdir. Düğüm noktasında yer alan yaygın örnekler arasında parazitler, mikroplar, ayrıştırıcılar, saprotroflar. tüketiciler ve avcılar sayılabilir.[7][8]
Trofik düzeyler
Besin ağlarında trofik düzeyler ve konumlar bulunur. İlk katmanda bulunan bitkiler gibi temel türler besin ağı içinde yer alan diğer türlerle beslenmeyen türlerdir. Temel türler ototrof ya da detritivor olabilir.[9] Ototrofların çoğu güneş enerjisini klorofil içinde tutar ancak litotrof bazı ototroflar biyoenerjiyi inorganik bileşiklerin kimyasal oksidasyonundan elde eder ve ışıksız ortamlarda yaşayabilirler; örneğin kükürt kaynaklarında yaşayan Thiobacillus cinsi bakteri gibi. En üst düzeyde bulunan süper avcılar besin ağı içerisinde başka bir tür tarafından besin olarak öldürülmeyen canlılardan oluşur. Ara düzeylerde ise birden fazla trofik düzeyden beslenen ve temel türlerden başlayarak çeşitli beslenme yollarından enerji akışını sağlayan hepçil türlerden oluşur.[10]
En basit anlamda besin ağının birinci trofik düzeyi bitkilerden, ikinci düzeyi otçullardan ve üçüncü düzeyi de etçillerden oluşur. Trofik düzey tepe tüketiciyi temel üreticiye bağlayan trofik bağ sayısından bir fazladır. Besin zincirinin temelinde yer alan ana üreticiler ya da detrivorlar "sıfır" düzeyi olarak kabul edilir.[1][11] Çevrebilimciler trofik türleri tanımlayabilmek için çeşitli türlerin midelerinde bulunan besin maddelerini yaygın olarak araştırarak beslenme bağlarını ortaya çıkarırlar. Bu teknik besin ağı içerisindeki enerji akışını daha iyi izleyebilmek için kararlı izotopların kullanılması yoluyla geliştirilmiştir.[12] Bir zamanlar hepçilliğin ender rastlandığı sanılsa da son zamanlardaki bulgular bunun terini göstermektedir ve bu bulgular trofik sınıflandırmaları daha karmaşık hâle getirmiştir.[13]
Trofik dinamiği
Trofik düzeyler kavramı Raymond L. Lindeman tarafından 1942 yılında yayımlanan blimsel bir makale ile ortaya çıkmıştır. Trofik dinamiğinin temeli ekosistemin bir kısmından diğer kısmına enerji transferidir.[11][14] Trofik dinamiği kavramı yaralı kantitatif buluşsal bir yöntemdir ancak bir organizmanın hangi spesifik trofik düzey içinde alınmasındaki hassasiyet de dahil olmak üzere önemli sınırlamaları vardır. Örneğin hepçiller tek bir trofik düzey ile sınırlandırılamamktadır. Yine de son zamanlarda yapılan araçtırmalar birbirinden ayrı trofik düzeylerin olduğunu ortaya çıkarmıştır ancak otçul trofik düzeyin üzerinde besin ağları hepçillerin girift bağlantıları ile tanımlanabilmektedir.[13]
Trofik dinamik literatürünün ana sorunsalı kaynaklar ve üretim üzerindeki kontrol ve düzenleme mekanizmasının doğasıdır. Çevrebilimciler üretici, etçil ve ayrıştırıcı düzeylerinden oluşan tek trofik konumlu basitleştirilmiş besin zinciri modelleri kullanmaktadırlar. Bu modelleri kullanarak çeşitli ekolojik kontrol mekanizmaları test edilmiştir. Örneğin geniş bitkisel kaynaklara sahip olan otçulların populasyonları genellikle avcılar tarafından kontrol altında tutulmakta ve düzenlenmektedir. Bu "yukarıdan aşağıya" ya da "yeşil dünya" varsayımı olarak bilinir. Alternatif olarak tüm bitkisel besinlerin yenilebilir olmadığı ve bitkisel besinlerin besin değeri ile bitkilerin otçullara karşı yapısal ya da kimyasal savunma mekanizmalarının güçlü olduğu durumlarda kontrol ve düzenleme mekanizması "aşağıdan yukarıya" bir yapıdadır.[15][16][17] Yakın zamanlarda yapılan araştırmalar hem "yukarıdan aşağıya" hem de "aşağıdan yukarıya" kuvvetlerin ekotopluluk yapısını etkilediğini ve etkinin gücünün çevresel koşullara bağlı olduğunu göstermiştir.[18][19] Bu karmaşık multitrofik etkileşimler besin ağında ikiden fazla trofik düzeyi kapsamaktadır.[20]
Multitrofik etkileşimlere bir başka örnek avcıların otçul populasyonunu kontrol altında tutarak bitkileri aşırı tüketmesini engellemesi ve bitki populasyonunun artışına yardım etmesi olan trofik zinciridir. Besin ağındaki bağlar türler arasındaki doğrudan trofik ilişkileri tasvir etse de trofik düzeyler arasında biyokütleyi, dağılımı ve bolluğu değiştirebilen dolaylı etkileri de vardır. Örneğin otçulları yiyen avcılar dolaylı olarak bitkilerin üretimini kontrol edip düzenlemektedir. Avcılar doğrudan bitkiler ile beslenmese de, bitki populasyonuna bağlı olan otçul populasyonunu düzenlemektedirler. Doğrudan ve dolaylu ilişkilerin net etkisi trofik zinciri olarak adlandırılır. Trofik zincirler besin ağı dinamiği içinde sınırlı bir populasyonu etkileyen tür düzeyinde zincir ile örneğin bitki biyokütlesinin dağılımı gibi besin ağının tamamı üzerinde oldukça dramatik etkileri olabilen topluluk düzeyi zincirler olarak ikiye ayrılır.[21]
Enerji akışı ve biyokütle
Besin ağları enerji akışını trofik bağlarla gösterir. Besin ağı sistemleri içinde döngüsel olan madde akışının aksine enerji akışı belirli bir yönde ilerler.[22] Enerji akışı tipik olarak üretimi, tüketimi, sindirimi, sindirilemeyen kayıpları (atıkları) ve solunumu (idame bedelini) içerir.[3] Genel olarak enerji akışı (E), metabolik üretim (P) ile solunumun (R) toplamı olarak yani E=P+R denklemiyle ifade edilebilir.
Herhangi bir şeyin kütlesi (ya da biyokütlesi) o şeyin enerji içeriğine denktir. Kütle ile enerji birbirleriyle içiçe geçmiştir. Ancak besinlerin konsantrasyonu ile kalitesi ve enerjisi değişkendir. Birçok bitki lifi çoğu otçul için sindirilebilir değildir dolayısıyla besin ve enerji kaynaklarına ulaşımı daha geniş olan bakterilerin bulunduğu atıksal besin ağlarına göre otçul besin ağı toplulukları besin açısından daha sınırlıdırlar.[23][24] Organizmalar genellikle enerjiyi karbonhidrat, lipit ve protein biçiminde özümserler. Bu polimerlerin hem enerji deposu hem de yapı taşları olarak çifte işlevi vardır. Enerji deposu olarak işlevleri besin ile birlikte karbondioksit, su ve ısı üretimi olarak sonuçlanır. Besin üretimi dolayısıyla metabolizma için temel bir işlevdir.[24] Enerji akışı ağlarında kullanılan birim tipik olarak birim zamanda m2'ye düşen kütle ya da enerji ölçüsüdür. Farklı tüketiciler beslenmelerinde farklı metabolik sindirme verimliliğine sahiptir. Her trofik düzey enerjiyi biyokütleye çevirir. Enerji akışı diyagramları bir trofik düzeyden diğerine, en tepe noktaya kadar iletim oranı ve verimliliğini tarif eder.[25][26]
Her trofik düzeyin biyokütlesi temelden tepeye doğru azalarak ilerler. Çünkü her enerji iletiminde çevreye doğru enerji kaybedilir ve entropi artar. Enerjinin yaklaşık yüzde seksen ile doksanı organizmanın yaşam süreçleri için harcanır ya da ısı ve atık olarak kaybedilir. Bir organizmanın enerjisinin yalnızca yüzde on kadar bir kısmı diğer organizmaya geçer.[27] Daha az sindirilebilir bitki tüketen hayvanlarda bu oran yüzde bire düşerken fitoplankton tüketen zooplanktonlarda ise yüzde kırka kadar çıkabilmektedir.[28] Her trofik düzeyde bulunan biyokütle ve verimliliğin grafiksel tasviri besin piramidi ya da ekolojk piramit olarak adlandırılır. Birincil üreticilerden tepe tüketicilere kadar olan enerji iletimi ayrıca enerji akış diyagramları ile de betimlenebilir.[29]
Besin zinciri
Besin ağı trofik yapısını ölçmek için yaygın olarak kullanılan ölçülerden birisi bezin zinciri uzunluğudur. Besin zinciri uzunluğu enerjinin ve besinin bitkilerden avcılara kadar hareket ettiği sırada karşılaştığı tür sayısını ölçüsü olarak besin ağını tarif etmenin başka bir yoludur.[30] Besin ağı dinamiği içinde enerji, etkileşim ve karmaşıklık gibi değişik parametrelere göre besin zinciri uzunluğu hesaplamanın farklı yolları vardır.[30] En basit hâliyle bir besin zincirinin uzunluğu trofik tüketici ile ağın tabanı arasında bulunan trofik bağ sayısıdır. Bir besin ağının ortalama besin zinciri uzunluğu ise ağ içinde bulunan tüm besin zincirlerinin aritmetik ortalamasıdır.[2][10]
Genellikle bir besin zincirinde dört ya da beş bağ bulunur ancak su ekosistemlerinde karaya nazaran daha uzun besin zincirleri gözlemlenir. Eninde sonunda besin zincirindeki tüm enerji ısı olarak çevreye dağılır.[2]
Basit bir avcı-av örneğinde bir geyik yediği bitkiden bir adım ötededir (zincir uzunluğu = 1); geyiği yiyen kurt ise bitkiden iki adım ötededir (zincir uzunluğu = 2). Bu parametrelerin besin ağı içindeki miktarı ve nüfuz gücü şu konular hakkındadır:
- Birkaç baskın türün varlığı ve tanımı (güçlü etkileşimciler ya da kilittaşı türler),
- Türlerin toplam sayısı ve besin zinciri uzunluğu (çok sayıda zayıf etkileşimciyi de içerir) ve
- Topluluğun yapısının, işlevinin ve kararlılığının nasıl belirlendiği[31][32]
Besin piramidi
Sayı besin piramidi tabanda üreticilerden başlayarak tepe noktada tek bir tüketiciye kadar her düzeyde tüketici sayısı azalarak oluşur. Besin piramitleri alan başına düşen birey sayısı, biyokütle (g/m2) ve enerji (k cal m−2 yr−1) gibi ekosistemin değişik sayısal özeliklerini göstermek için kullanılır. ALt trofik düzeylerden üst trofik düzeylere çıktıkça iletilen enerji miktarının azalması tüm ekosistemlerde gözlemlenen yapılardan biridir.[1][33][34] Piramidin her düzeyinin boyutu genellikle bir organizmanın kuru ağırlığı olark ölçülen biyokütledir.[35] Ototroflar dünya üzerinde en büyük biyokütle oranına sahiptir ancak mikropların biyokütlesi ototroflara çok yakındır ve hatta daha da fazla olabilir.[36][37]
Piramidin yapısı ekosistemden ekosisteme ve zamana göre değişiklik gösterebilir. Bazı durumlarda besin piramidi yapısı tersine dönebilir; bu yapıya su ve mercan kayalığı ekosistemlerinde rastlanır. Biyokütlenin tersine dönmüş yapısı genellikle üreticilerin boyutları ile alakalıdır. Suda yaşayan topluluklarda genellikle üreticiler, yüksek büyüme hızına sahip tüketicilerden daha küçük boyutludurlar. Planktonik alglar ya da su bitkileri gibi üreticilerde kara ekosistemlerindeki bitkilerde görülen sekonder büyüme görülmez. Ancak daha büyük bir otçul populasyonu besleyebilmek için daha hızlı üreyebilirler. Bu nedenle su sistemlerindeki piramit tersine döner. Ana tüketiciler, tükettikleri üreticilere göre daha uzun yaşam süresine ve daha yavaş büyüme hızına sahiptirler. Fitoplankton yalnızca bir iki yaşarken, fitoplankton ile beslene zooplankton birkaç hafta hayatta kalır ve zooplankton ile beslenen balıklar ise birkaç yıl yaşar.[38] Su avcılarının ölüm oranı da daha küçük tüketicilere göre daha düşüktür bu da piramidin tersine dönmesine yardımcı olur. Populasyon yapısı, göç oranları ve çevrede avcılardan saklanma olanakları tersine dönmüş biyokütle piramidinin diğer olası nedenleridir. Ancak enerji piramitleri, tüm besin enerji kaynakları dahil edildiğinde termodinamiğin ikinci yasası gereği her zaman doğru piramit şeklindedir.[2][39]
Madde akışı ve geri dönüşüm
Dünya üzerinde bulunan element ve minerallerin çoğu organizmaların dokularında ve aldıkları besinlerde bulunur. Bu nedenle mineral ve besin maddeleri döngüleri besin ağı enerji iletim yollarını izlerler. Çevrebilimciler tüm organizmalarda bulunan karbon (C), azot (N) ve fosfor (P) gibi ana elementlerin oranlarını analiz etmek için stokiyometri kullanırlar. Karasal ve su ekosistemleri arasında büyük geçiş farklılıkları bulunur; karasal sistemlerde C:P ve C:N oranları daha fazla iken N:P oranları her iki ekosistemde de aynıdır.[40][41][42] Mineral besin kaynakları organizmaların büyüme, gelişme ve hayatta kalma için gereksinim duyduğu maddesel kaynaklardır. Besin ağları mineral besinlerin organizmalar arasında geçişlerini betimler.[2][14] Ekosistemlerdeki ana üretimin çoğu tüketilmez ancak ayrıştırıcılar tarafından tekrar yararlı besinlere dönüştürülür.[43] Dünya üzerindeki mikroorganizmaların çoğu biyomineralizasyon adı verilen süreç içinde yer alarak minerallerin oluşmasını sağlar.[44][45][46] Atık ya da ölü organik madde sedimentlerinde yaşayan bakteriler besin maddelerini ve biyomineralleri yaratarak dönüştürürler.[47] Besin ağı ve madde döngüsü modelleri geleneksel olarak ayrı ayrı ele alınmış olsa da birbirleri arasında kararlılık, akış, kaynaklar, ve mineral besinleri geri dönüşümü gibi gşçlü işlevsel bağlar bulunur.[48][49]
Kaynakça
- 1 2 3 Pimm, S. L.; Lawton, J. H.; Cohen, J. E. (1991). "Food web patterns and their consequences" (PDF). Nature 350 (6320): 669–674. DOI:10.1038/350669a0. http://www.nicholas.duke.edu/people/faculty/pimm/publications/pimmreprints/71_Pimm_Lawton_Cohen_Nature.pdf.
- 1 2 3 4 5 Odum, E. P.; Barrett, G. W. (2005). Fundamentals of ecology. Brooks Cole. s. 598. ISBN 978-0-534-42066-6. http://www.cengage.com/search/totalsearchresults.do?N=16&image.x=0&image.y=0&keyword_all=fundamentals+of+ecology.
- 1 2 Benke, A. C. (2010). "Secondary production". Nature Education Knowledge 1 (8): 5. http://www.nature.com/scitable/knowledge/library/secondary-production-13234142.
- ↑ Allesina, S.; Alonso, D.; Pascual, M.. "A general model for food web structure.". Science 320 (5876): 658–661. DOI:10.1126/science.1156269. http://cas.bellarmine.edu/tietjen/Secret/PlantGenome/General%20Model%20for%20Food%20WEb%20Structure.pdf.
- ↑ Azam, F.; Fenche, T.; Field, J. G.; Gra, J. S.; Meyer-Reil, L. A.; Thingstad, F. (1983). "The ecological role of water-column microbes in the sea". Mar. Ecol. Prog. Ser. 10: 257–263. DOI:10.3354/meps010257. http://www.soest.hawaii.edu/oceanography/courses/OCN621/Spring2011/Azam%20et%20al_loop.pdf.
- ↑ Uroz, S.; Calvarus, C.; Turpault, M.; Frey-Klett, P. (2009). "Mineral weathering by bacteria: ecology, actors and mechanisms". Trends in Microbiology 17 (8): 378–387. DOI:10.1016/j.tim.2009.05.004. PMID 19660952. http://mycor.nancy.inra.fr/GIteam/wp-content/uploads/2009/11/Uroz-TiM-2009.pdf.
- ↑ Williams, R. J.; Martinez, N. D. (2000). "Simple rules yield complex food webs.". Nature 404 (6774): 180–183. DOI:10.1038/35004572. http://userwww.sfsu.edu/~parker/bio840/pdfs/WilliamsMartinez2000.pdf.
- ↑ Post, D. M. (2002). "The long and short of food chain length". Trends in Ecology and Evolution 17 (6): 269–277. DOI:10.1016/S0169-5347(02)02455-2. http://limnology.wisc.edu/courses/zoo955/Spring2005/food%20web%20seminar%20papers/post02TREE.pdf.
- ↑ Tavares-Cromar, A. F.; Williams, D. D. (1996). "The importance of temporal resolution in food web analysis: Evidence from a detritus-based stream". Ecological Monographs 66 (1): 91–113. DOI:10.2307/2963482.
- 1 2 Pimm, S. L. (1979). "The structure of food webs". Theoretical population biology 16: 144–158. DOI:10.1016/0040-5809(79)90010-8. PMID 538731. http://www.nicholas.duke.edu/people/faculty/pimm/publications/pimmreprints/12_Pimm_TPB_1979.pdf.
- 1 2 Cousins, S.. "Ecologists build pyramids again.". New Scientist 1463: 50–54. https://books.google.com/books?id=NOPpwVvNu44C&pg=PA51&dq=trophic+level#v=onepage&q=trophic%20level&f=false.
- ↑ McCann, K. (2007). "Protecting biostructure". Nature 446 (7131): 29. DOI:10.1038/446029a. PMID 17330028. http://www.bolinfonet.org/pdf/McCann_2007_biostructure.pdf.
- 1 2 Thompson, R. M.; Hemberg, M.; Starzomski, B. M.; Shurin, J. B. (March 2007). "Trophic levels and trophic tangles: The prevalence of omnivory in real food webs.". Ecology 88: 612–617. DOI:10.1890/05-1454. PMID 17503589. http://myweb.dal.ca/br238551/thompson_hem_star_shur_ecology07.pdf.
- 1 2 Lindeman, R. L. (1942). "The trophic-dynamic aspect of ecology". Ecology 23 (4): 399–417. DOI:10.2307/1930126. http://www.fcnym.unlp.edu.ar/catedras/ecocomunidades/Lindman_1942.pdf.
- ↑ Hairston, N. G.; Hairston, N. G. (1993). "Cause-effect relationships in energy flow, trophic structure, and interspecific interactions.". The American Naturalist 142 (3): 379–411. DOI:10.1086/285546. http://limnology.wisc.edu/courses/zoo955/Spring2005/food%20web%20seminar%20papers/hairston93AmNat.pdf.
- ↑ Fretwell, S. D. (1987). "Food chain dynamics: The central theory of ecology?". Oikos 50: 291–301. DOI:10.2307/3565489. http://limnology.wisc.edu/courses/zoo955/Spring2005/food%20web%20seminar%20papers/fretwell_food_chain_dynamics_oikos.pdf.
- ↑ Polis, G. A.; Strong, D. R. (1996). "Food web complexity and community dynamics.". The American Naturalist 147 (5): 813–846. DOI:10.1086/285880. http://www.seaturtle.org/PDF/Polis_1996_AmerNat.pdf.
- ↑ Hoekman, D.. "Turning up the head: Temperature influences the relative importance of top-down and bottom-up effects.". Ecology 91 (10): 2819–2825. DOI:10.1890/10-0260.1. http://www.nd.edu/~underc/east/publications/documents/Hoekman2010b.pdf.
- ↑ Schmitz, O. J. (2008). "Herbivory from individuals to ecosystems.". Annual Review of Ecology, Evolution, and Systematics 39: 133–152. DOI:10.1146/annurev.ecolsys.39.110707.173418. http://www.annualreviews.org/doi/abs/10.1146/annurev.ecolsys.39.110707.173418.
- ↑ Tscharntke, T.; Hawkins, B., A., ed. (2002). Multitrophic Level Interactions. Cambridge: Cambridge University Press. s. 282. ISBN 0-521-79110-3. https://books.google.com/books?id=_8bHeXvUc08C&printsec=frontcover&dq=Multitrophic+Level+Interactions#v=onepage&q&f=false.
- ↑ Polis, G.A. (2000). "When is a trophic cascade a trophic cascade?". Trends in Ecology and Evolution 15 (11): 473–5. DOI:10.1016/S0169-5347(00)01971-6. PMID 11050351. http://www.cof.orst.edu/leopold/class-reading/Polis%202000.pdf.
- ↑ Odum, E. P. (1968). "Energy flow in ecosystems: A historical review". American Zoologist 8 (1): 11–18. DOI:10.1093/icb/8.1.11. http://icb.oxfordjournals.org/content/8/1/11.short.
- ↑ Mann, K. H. (1988). "Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems". Limnol. Oceanogr. 33 (2): 910–930. DOI:10.4319/lo.1988.33.4_part_2.0910. http://nospam.aslo.org/lo/toc/vol_33/issue_4_part_2/0910.pdf.
- 1 2 Koijman, S. A. L. M.; Andersen, T.; Koo, B. W. (2004). "Dynamic energy budget representations of stoichiometric constraints on population dynamics". Ecology 85 (5): 1230–1243. DOI:10.1890/02-0250. http://www.bio.vu.nl/thb/research/bib/KooyAnde2004.pdf.
- ↑ Anderson, K. H.; Beyer, J. E.; Lundberg, P. (2009). "Trophic and individual efficiencies of size-structured communities". Proc Biol Sci. 276 (1654): 109–114. DOI:10.1098/rspb.2008.0951. PMC 2614255. PMID 18782750. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2614255.
- ↑ Benke, A. C. (2011). "Secondary production, quantitative food webs, and trophic position". Nature Education Knowledge 2 (2): 2. http://www.nature.com/scitable/knowledge/library/secondary-production-quantitative-food-webs-and-trophic-17653963.
- ↑ Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. s. 165. ISBN 978-1-4200-5544-3. https://books.google.com/?id=Grivqd7tLuAC&pg=PA165&dq=%22is+lost+as+heat+and+wastes%22&cd=2#v=onepage&q=%22is%20lost%20as%20heat%20and%20wastes%22.
- ↑ Kent, Michael (2000). Advanced Biology. Oxford University Press US. s. 511. ISBN 978-0-19-914195-1. https://books.google.com/?id=8aw4ZWLABQkC&pg=PA511&dq=%22trophic+efficiency+of+less+than+1%25%22&cd=1#v=onepage&q=%22trophic%20efficiency%20of%20less%20than%201%25%22.
- ↑ Kent, Michael (2000). Advanced Biology. Oxford University Press US. s. 510. ISBN 978-0-19-914195-1. https://books.google.com/?id=8aw4ZWLABQkC&pg=PA510&dq=%22by+an+energy+flow+diagram%22&cd=1#v=onepage&q=%22by%20an%20energy%20flow%20diagram%22.
- 1 2 Post, D. M. (1993). "The long and short of food-chain length". Trends in Ecology and Evolution 17 (6): 269–277. DOI:10.1016/S0169-5347(02)02455-2.
- ↑ Worm, B.; Duffy, J.E. (2003). "Biodiversity, productivity and stability in real food webs". Trends in Ecology and Evolution 18 (12): 628–632. DOI:10.1016/j.tree.2003.09.003.
- ↑ Paine, R. T. (1980). "Food webs: Linkage, interaction strength and community infrastructure.". Journal of Animal Ecology 49 (3): 666–685. DOI:10.2307/4220. JSTOR 4220.
- ↑ Proulx, Stephen R.; Promislow, Daniel E.L.; Phillips, Patrick C. (2005). "Network thinking in ecology and evolution". Trends in Ecology and Evolution 20 (6): 345–353. DOI:10.1016/j.tree.2005.04.004. PMID 16701391.
- ↑ Raffaelli, D. (2002). "From Elton to mathematics and back again". Science 296 (5570): 1035–1037. DOI:10.1126/science.1072080. PMID 12004106.
- ↑ Rickleffs, Robert, E. (1996). The Economy of Nature. University of Chicago Press. s. 678. ISBN 0-7167-3847-3.
- ↑ Whitman, W. B.; Coleman, D. C.; Wieb, W. J. (1998). "Prokaryotes: The unseen majority". Proc. Natl. Acad. Sci. USA 95 (12): 6578–83. DOI:10.1073/pnas.95.12.6578. PMC 33863. PMID 9618454. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=33863.
- ↑ Groombridge, B.; Jenkins, M. (2002). World Atlas of Biodiversity: Earth's Living Resources in the 21st Century. World Conservation Monitoring Centre, United Nations Environment Programme. ISBN 0-520-23668-8. https://books.google.com/?id=_kHeAXV5-XwC&printsec=frontcover.
- ↑ Spellman, Frank R. (2008). The Science of Water: Concepts and Applications. CRC Press. s. 167. ISBN 978-1-4200-5544-3. https://books.google.com/?id=Grivqd7tLuAC&pg=PA167&dq=However,+biomass+pyramids+can+sometimes+be+inverted.&cd=1#v=onepage&q=However%2C%20biomass%20pyramids%20can%20sometimes%20be%20inverted..
- ↑ Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). "Modeling inverted biomass pyramids and refuges in ecosystems". Ecological Modelling 220 (11): 1376–1382. DOI:10.1016/j.ecolmodel.2009.03.005. http://people.math.gatech.edu/~weiss/pub/General_Mechanisms_Final.pdf.
- ↑ Pomeroy, L. R.. "The strategy of mineral cycling". Annual Review of Ecology and Systematics 1: 171–190. DOI:10.1146/annurev.es.01.110170.001131. JSTOR 2096770.
- ↑ Elser, J. J.; Fagan, W. F.; Donno, R. F.; Dobberfuhl, D. R.; Folarin, A.; Huberty, A. (2000). "Nutritional constraints in terrestrial and freshwater food webs". Nature 408 (6812): 578–580. DOI:10.1038/35046058. http://www.rug.nl/biologie/onderzoek/onderzoekInstituten/cees/colloquia/pdf/elser_et_al2_2000.pdf.
- ↑ Koch, P. L.; Fox-Dobbs, K.; Newsom, S. D.. Diet, G. P.; Flessa, K. W.. ed. The isotopic ecology of fossil vertebrates and conservation paleobiology.. 15. s. 95–112. http://www.es.ucsc.edu/~pkoch/pdfs/Koch%20papers/2009/Koch%20et%2009%20PSP%2015.pdf.
- ↑ Moore, J. C.; Berlow, E. L.; Coleman, D. C.; de Ruiter, P. C.; Dong, Q.; Hastings, A. (2004). "Detritus, trophic dynamics and biodiversity". Ecology Letters 7 (7): 584–600. DOI:10.1111/j.1461-0248.2004.00606.x.
- ↑ H. A., Lowenstam (1981). "Minerals formed by organisms.". Science 211 (4487): 1126–1131. DOI:10.1126/science.7008198. JSTOR 1685216. PMID 7008198.
- ↑ Warren, L. A.; Kauffman, M. E. (2003). "Microbial geoengineers". Science 299 (5609): 1027–1029. DOI:10.1126/science.1072076. JSTOR 3833546. PMID 12586932.
- ↑ González-Muñoz, M. T.; Rodriguez-Navarro, C.; Martínez-Ruiz, F.; Arias, J. M.; Merroun, M. L.; Rodriguez-Gallego, M.. "Bacterial biomineralization: new insights from Myxococcus-induced mineral precipitation.". Geological Society, London, Special Publications 336 (1): 31–50. DOI:10.1144/SP336.3. http://sp.lyellcollection.org/content/336/1/31.abstract.
- ↑ Gonzalez-Acosta, B.; Bashan, Y.; Hernandez-Saavedra, N. Y.; Ascencio, F.; De la Cruz-Agüero, G.. "Seasonal seawater temperature as the major determinant for populations of culturable bacteria in the sediments of an intact mangrove in an arid region.". FEMS Microbiology Ecology 55 (2): 311–321. DOI:10.1111/j.1574-6941.2005.00019.x. http://www.bashanfoundation.org/gmaweb/pdfs/mangrovebalandra.pdf.
- ↑ DeAngelis, D. L.; Mulholland, P. J.; Palumbo, A. V.; Steinman, A. D.; Huston, M. A.; Elwood, J. W. (1989). "Nutrient dynamics and food-web stability.". Annual Review of Ecology and Systematics 20: 71–95. DOI:10.1146/annurev.ecolsys.20.1.71. JSTOR 2097085.
- ↑ Twiss, M. R.; Campbell, P. G. C.; Auclair, J. (1996). "Regeneration, recycling, and trophic transfer of trace metals by microbial food-web organisms in the pelagic surface waters of Lake Erie.". Limnology and Oceanography 41 (7): 1425–1437. DOI:10.4319/lo.1996.41.7.1425. http://www.nospam.aslo.org/lo/toc/vol_41/issue_7/1425.pdf.