Uçbirleştirme (genetik)

Genetikte uçbirleştirme (İngilizce splicing) transkripsiyon sonrasında RNA'daki bazı bölümlerin (intron'ların) çıkartılıp kalan kısımlarının (eksonların) birleştirilmesidir. Ökaryotlarda uçbirleştirme sonucunda prekürsör mesajcı RNA (pre-mRNA) ergin mesajcı RNA'ya dönüşür, bu da protein sentezinde kullanılır. Uçbirleştirme bir dizi biyokimyasal tepkimeden oluşur, bunlar splisozom adı verilen bir snRNP (İngilizce small nuclear ribonucleo-proteins, küçük çekirdeksel ribonükleoproteinler) kompleksi tarafından katalizlenir.
RNA'dan parçaların çıkarılıp kalan kısımların uçlarının birleştirilmesi süreci 1977'de Philip Sharp ve Richard J. Roberts tarafından bağımsız olarak keşfedildi.[1][2] Sharp ve Roberts bu kesiflerinden dolayı 1993'de Nobel Fizyoloji veya Tıp Ödülünü kazandılar.
Etimoloji
RNA'dan parçaların çıkarılıp kalan kısımların birleştirilmesi sürecini tarif etmek için kullanılan İngilizce sözcük, splicing ("splaysing" okunur; Almanca splissen 'den türeme), orijinal anlamıyla bir denizcilik terimi olup, kopmuş bir halatı tamir etmek amacıyla iki ucunu birleştirme işlemi (halat dikişi) için kullanılır. Bu denizcilik terimi, zaman içinde kalas, demiryolu rayı gibi başka cisimlerin uçlarının birleştirilmesi için, ve 20 y.y.'da sinemacılıkta kopan filmin tamiri ve elektrik kablo birleştirmesi için kullanılmıştır (bkz. uç birleştirme (anlam ayrımı)). RNA'da ekson ve intronların keşfi üzerine bu terime ekson uçlarının birleştirilmesi kavramı da yüklenmiştir.[3] Bu biyolojik olguya Fransızca ve Almanca'da bulunmuş karşılıklar, épissage ve spleißen de aynı sekilde, genel kapsamlı "uç birleştirme" anlamını taşımaktadır. Splicing için Türkçede karşılık henüz yerleşmediği için, buradaki metinde, aynı anlam yükleme süreci uygulanarak "uçbirleştirme" terimi kullanılmıştır.
Uçbirleştirme yolları
Doğada RNA uç birleştirmesi birkaç yolla gerçekleşir. Uç birleştirmenin tipi çıkartılacak intronun yapısı ve bu tepkime için gereken katalizöre bağlıdır. Hangi yol kullanılırsa kullanılsın, çıkartılan intron kullanılmadan yıkıma uğrar.
Splisozomal intronlar
Splisozomal intronlar genelde ökaryotların protein kodlayıcı genlerinde bulunurlar. Uç birleşme için intronun içinde bir 3' birleştirme ucu, bir 5' birleştirme ucu, bir de bir dal noktası gereklidir. Uç birleştirme splisozom tarafından katalizlenir, bu, beş ribonükleoproteinden (snRNP'den; "snörp" diye okunur) oluşan bir RNA-protein kompleksidir. snRNP'lerdeki RNA'lar belli intron dizileri ile etkileşirler ve uçbirleşmenini katalizinde rol oynayabilirler. İki farklı tip splisozom bilinmektedir, esas (majör) ve ikincil (minör) splisozomlar farklı snRNP'lar içerir.
- Majör splisozomlar
- Majör splisozom, 5' uç birleşme konumunda GU ve 3' uç birleşme konumunda AG dizisi olan intronların uç birleştirmesinde rol oynar. Onu oluşturan snRNP'ler U1, U2, U4, U5, and U6 snRNP olarak adlandırılır. Bu tip uçbirleştirmeye "standart uçbirleştirme" (İng. canonical splicing) veya "kement yolu" (İng. lariat pathway) adı verilir, uçbirleştirmelerin %99'undan çoğunu kapsar. Buna karşın, intron uçlarındaki diziler GU-AG kuralına uymazsa bunun "standart dışı" uçbirleştirme (İng. noncanonical splicing) olduğu söylenir (bkz. aşağıda "minör splisozomlar").[4]
- Minör splisozomlar
- Minör splisozom majör splisozoma çok benzer, ancak uçbirleşme dizileri farklı olan ender intronları çıkartır. Hem minör hem majör splisozomda U5 snRNP bulunmasına karşın, minör splisozomda U1, U2, U4, ve U6 snRNP'ye benzer işlevleri olan U11, U12, U4atac, ve U6atac adlı farklı snRNP'ler bulunur.[5]
- Trans-uçbirleştirme
- Trans-uçbirleştirme aynı RNA transkriptinin içinde yer almayan iki eksonun birleştirilmesidir. Splisozom aracılığıyla iki farklı prekürsör RNA'daki eksonlar birbirleriyle birleşirler. Trans-uçbirleştirme protozoalrda (tripanozomlar ve öglenoidlerde), sineklerde (Drosophila) ve solucan Caenorhabditis elegans 'ta görülmüştür.
Öz-uçbirleştirme
Öz-uçbirleştirme (İngilizce self-splicing), veya otokatalitik uçbirleştirme, ribozim oluşturabilen ender intronlarda meydana gelir, bunlarda splisozomun katalizörlük işlevi yalnızca RNA tarafından sağlanır. İki cins öz-uçbirleştiren intron vardır, bunlar Grup I ve Grup II katalitik intron diye adlandırılır. Grup I ve II intronların kullandığı mekanizma bir protein varlığını gerektirmese de uçbirleştirmenin yanlışsız olabilmesi için ATP gereklidir. Bu ATP'nin üretilmesi için çeşitli metabolizma enzimleri gereklidir. Grup I intronlu uçbirleştirmede iki transesterifikasyon tepkimesi vardır: 1) GDP, GTP veya GMP gibi bir kofaktörde yer alan bir guanin nükleozitin 3' OH grubu, 5' uçbirleşme konumundaki fosfata bağlanır. 2) İntronun 5' ucundaki 3' OH grubu bir nükleofil olur, ikinci bir transesterifikasyon ile iki ekson birleşir. İntron, lineer bir parça olarak salınır. Grup II intronlu uçbirleştirmenin mekanizması ise şöyledir: 1) İntrondaki belli bir adenozinin '2 OH grubu, 5' uçbirleştirme konumuna saldırır, böylece "kement" (lariat) oluşturur. 2) Eksonun 5' ucundaki nukleotidin 3' OH grubu, 3' uçbirletirme konumumu ile ikinci bir transesterifikasyonu tetikler, böylece iki ekson birleşir, intron ise kement biçimli bir parça olarak salınır.
tRNA uçbirleştirmesi
tRNA uçbirleştirmesi, veya tRNA-benzeri uçbirleştirme, genelde tRNA'da meydana gelen ender bir uçbirleştirme biçimidir.
Uçbirleştirme tepkimesinin mekanizması splisozomal ve öz-uçbirleştirme yollarından farklıdır. RNA, ribonükleaz tarafından kesilir, eksonlar ligazlar tarafından birleştirilir. Bu tip uçbirleştirmenin katalizlenmesi için herhangi bir RNA unsur gerekli değildir.
Evrim
Uçbirleştirme canlılardaki tüm alem veya üst alemlerde görülür, ancak bu gruplar içinde uçbirleştirmenin yaygınlığı ve tipi çok fark gösterebilir. Ökaryotlar pek çok protein kodlayıcı mesajcı RNA ve bazı kodlamayıcı RNA'yı uçbirleştirir. Buna karşın prokaryotlar, ender olarak, ve başlıca kodlamayıcı RNA'yı uçbirleştirir. Bu iki grup organizma arasındaki önemli bir diğer fark, prokaryotlarda splisozom yolunun bulunmamasıdır.
| Ökaryotlar | Prokaryotlar | |
|---|---|---|
| Splisozomal | + | - |
| Öz-uçbirleştirme | + | + |
| tRNA | + | + |
Grup II intronların yapısı spisozomun içinde bulunan snRNP'lerin yapısına çok benzediği için splisozomların Grup II intronlardan evrimleşmiş olabileceğini düşünülmektedir.[6]
Biyokimyasal mekanizma
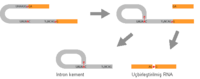
Splisozomal uçbirleştirme ve öz-uçbirleştirme de iki aşamalı bir biyokimyasal süreçtir. Her aşama, RNA nükleotidleri arasında oluşan transesterifikasyon tepkimelerinden oluşur. tRNA uçbirleştirmesi farklıdır, transesterfikasyon ile meydana gelmez.
Splisozomal ve öz-uçbirleştirme transterifikasyon tepkimeleri biribirini izleyen iki reaksiyondan oluşur. Önce, splisozomun bir araya gelmesi sonucu konumu belirlenen spesifik bir "dal noktası" nükleotidinin 2' OH grubu, intronun 5' uçbirleştirme konumundaki ilk nükleotid üzerine bir nükleofilik saldırı yaparak "kement ara ürünü"nü oluşturur. İkinci aşamada, serbest kalmış 5' eksonun 3' OH grubu, intronun 3' uçbirleştirme konumundaki son nükleotide bir nükleofilik saldırı yapar, böylece iki ekson birleşir, kementli intron da salınır.
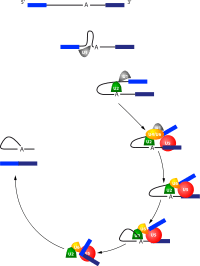
Splisozomal uçbirleştirmede snRNP'ler bu reaksiyonların gerçekleşmesini mümkün kılar. Bu süreç sırasında çeşitli snRNP'ler belli bir sıra içinde bir araya gelirler veya ayrışırlar:
- U1 intronun 5' ucuna bağlanır.
- U2 dallanma noktasına bağlanır.
- U4/U6 ikilisi U1 ve U2'ye yanaşarak intronun 5' ucu ile dallanma noktası arasında bir köprü kurarlar.
- U5 de komplekse katılır ve birleştirirlecek eksonların uçlarını bitiştirir.
- U4 ve U1 kompleksden ayrılırlar.
- Dallanma noktasındaki adenozinin 2'-OH grubu, intronun 5' ucunu keser.
- İntronun yukarı tarafındaki eksonun 3' ucundaki nükleotidin 3'-OH grubu, intronun öbür ucunu keser.
- Eksonların uçları birleşir ve intronun "kement" şekilli kalıntısı salınır.
SnRNP'ların RNA'ya bağlanması tümleyici nükleotit dizileri arasında baz ciftleşmleriyle gerşkleşir. Örneğin, snRNA U1, intronun 5' tarafındaki ekson-intron ekleminde bulunan konsensus dizisini tümleyici bir diziye sahiptir, bunun sayesinde RNA'nın orasına bağlanabilir. Ayrıca farklı snRNA'ların birbirlerine bağlanmalarını sağlayan RNA-RNA birleşme yerleri de mevcuttur.
Alternatif uçbirleştirme
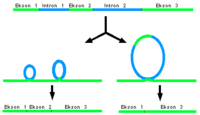
Çoğu genin uçbirleştirmesinde, bir ekson birden fazla sayıda eksondan biri ile birleşebilir. Mesajcı RNA'nın ekson bileşiminin bu şekilde değişmesi sonucunda bir dizi farklı protein oluşur. Bu olguya alternatif uçbirleştirme denir.
Uçbirleştirmenin deneysel manipülasyonu
Uçbirleştirme tepkimesindeki aşamalar deneysel olarak değişirilebilir. Bu amaçla, snRNP bağlanma konumlarını[7], kementi kapatan dal noktası nükleotidi[8] veya uçbirleştirme düzenleyici eleman bağlanma konumlarını[9] bloke edecek ters-anlamlı (İng. anti-sense) oligonükleotit veya oligonükleotit türevleri kullanılabilir.
Uçbirleştirme hataları
İntron ve eksonlarda mutasyonlar uçbirleştirmeyi engelleyebilir ve sonuç olarak kodlanan proteinin sentezini durdurabilirler.
Olabilecek mutasyonlar:
- Uçbirleştirme konumunda bir mutasyon, o yerin işlevini kaybetmesine neden olur. Bunun sonucunda ekson kaybı, intron içlemesi, veya erken stop kodonu oluşumu meydana gelebilir.
- Bazı mutasyonlar uçbirleştirmede spesifisite kaybına yol açar. Uçbirleştirme konumunda değişime neden olarak proteinde amino asit katılma veya eksilmeleri, veya daha muhtemel olarak, okuma çerçevesi (İng. reading frame) kaybı meydana gelir.
- Transpozisyon sonucu DNA kaybı veya eklenmesi meydana gelebilir, bu da normalden daha uzun veya daha kısa eksonlara yol açabilir.
Proteinlerde uçbirleştirme
Proteinlerde de uçbirleştirme olabilir. Bunun biyokimyasal mekanizması RNA uçbirleştirmesinden tamamen farklıdır. Proteinin intein olarak adlandırılan belli bir bölümü, çıkartılır; ekstein olarak adlandırılan proteinin kalan kısımları birleşir. Protein uçbirleştirmesi ekmek mayasında gözlemlenmiştir.
Notlar
- ↑ Spliced segments at the 5' terminus of adenovirus 2 late mRNA. Berget SM, Moore C, Sharp PA. Proc Natl Acad Sci U S A. 74(8):3171-5 1977
- ↑ An amazing sequence arrangement at the 5' ends of adenovirus 2 messenger RNA. Chow LT, Gelinas RE, Broker TR, Roberts RJ. Cell. 12:1-8.1977
- ↑ "Dictionary.com "splicing" tanımı". 21 Eylül 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150921013131/http://dictionary.reference.com:80/browse/splicing. Erişim tarihi: 2008-03-20.
- ↑ Ng B, Yang F, Huston DP, et al (Dec 2004). "Increased noncanonical splicing of autoantigen transcripts provides the structural basis for expression of untolerized epitopes". J. Allergy Clin. Immunol. 114 (6): 1463-70. Hata: Kötü DOI belirtildi. PMID 15577853.
- ↑ Patel AA, Steitz JA (2003). "Splicing double: insights from the second spliceosome". Nat. Rev. Mol. Cell Biol. 4 (12): 960-70. Hata: Kötü DOI belirtildi. PMID 14685174.
- ↑ P. A. Sharp, 1991, Science 254:663
- ↑ Draper BW, Morcos PA, Kimmel CB (2001). "Inhibition of zebrafish fgf8 pre-mRNA splicing with morpholino oligos: a quantifiable method for gene knockdown". Genesis 30 (3): 154-6. Hata: Kötü DOI belirtildi. PMID 11477696.
Sazani P, Kang SH, Maier MA, et al (Oct 2001). "Nuclear antisense effects of neutral, anionic and cationic oligonucleotide analogs". Nucleic Acids Res. 29 (19): 3965-74. PMID 11574678. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=60237. - ↑ Morcos, PA (2007). "Achieving targeted and quantifiable alteration of mRNA splicing with Morpholino oligos.". Biochem Biophys Res Commun 358 (2): 521-7. Hata: Kötü DOI belirtildi. PMID 17493584.
- ↑ Bruno IG, Jin W, Cote GJ (2004 Oct 15). "Correction of aberrant FGFR1 alternative RNA splicing through targeting of intronic regulatory elements". Hum. Mol. Genet. 13 (20): 2409-20. Hata: Kötü DOI belirtildi. PMID 15333583. http://hmg.oxfordjournals.org/cgi/content/full/13/20/2409.(Epub August 27, 2004)
Kaynakça
- Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James E. (1999). Molecular Cell Biology. New York: W. H. Freeman & Co.. ISBN 0-7167-3706-X.
- Hartl, Daniel L.; Elizabeth W. Jones (2005). Genetics: Analysis of Genes and Genomes. Jones & Bartlett Publishers. ISBN 0763715115.
Dış bağlantılar
- Omurgalı hayvanların pre-mRNA'larının 5' ve 3' uçbirleştirme konumlarındaki konsensus dizileri
- Grup I, Grup II öz-uçbirleştirme ve splisozomlu özbirleştirmelerinin biyokimyasal mekanizma şemaları
- Splizomu döngüsü.
- Grup II intronlar ve splsozomdaki snRNA'ların benzerliği
- Majör, minör ve trans-uçbirleştirme mekanzimaları