Fil
 Afrika fili (Loxodonta africana) | ||||||||||
| Bilimsel sınıflandırma | ||||||||||
| ||||||||||
| | ||||||||||
| Dış bağlantılar | ||||||||||
|
Fil, hortumlular takımının filgiller (Elephantidae) familyasını oluşturan memeli bir hayvandır. Geleneksel olarak Asya fili (Elephas maximus) ve Afrika fili (Loxodonta africana) olmak üzere iki türü tanınır; ancak bazı kanıtlara dayanarak Afrika savan fili (L. africana) ile Afrika orman filinin (L. cyclotis) de iki ayrı tür olduğu öne sürülür. Filler, Sahra altı Afrika ile Güney ve Güneydoğu Asya'da bulunur. İçinde mamutlar ve mastodonlar gibi soyu tükenmiş türleri de barındıran hortumlular takımından günümüzde soyunu sürdüren bir tek filler kalmıştır. Karada yaşayan en büyük hayvan olan Afrika filinin erkeği 4 m boya ve 7.000 kg ağırlığa ulaşabilir. Fillerin dikkat çekici ve ayırt edici özellikleri arasında, nesneleri yakalamak gibi çeşitli amaçlar için kullanılan uzun hortumları başta gelir. Çok uzun ve sivri olan kesici dişlerini nesneleri taşımak, yeri kazmak için kullanırlar. Fildişinin kaynağı olan bu kesici dişler aynı zamanda dövüşürken silah olarak da kullanılır. Filin büyük ve geniş kulakları vücut ısısını kontrol etmeye yarar. Afrika fillerinin kulakları daha büyük olur ve sırtları içbükeydir. Asya fillerinin ise kulakları daha küçük olur ve sırtları dışbükey ya da düzdür.
Otçul olan filler; savan, orman, çöl ve bataklık gibi doğal yaşam alanlarında bulunur. Genellikle su kenarlarında kalmayı tercih eder. Çevrelerinde bıraktıkları etki yüzünden kilittaşı türlerden biri sayılır. Diğer hayvanlar fillerden uzak durur ve aslan, kaplan, sırtlan ile yabani köpekler gibi yırtıcılar yalnızca yavru fillere saldırır. Dişi filler, genellikle bir dişi ve yavruları ya da akraba dişiler ve yavrularından oluşan aile grupları hâlinde yaşar. Birkaç dişi ve yavrularından oluşan grubun lideri en yaşlı dişidir. Fillerin oluşturduğu aile grupları zaman zaman bir araya gelerek daha büyük topluluklar oluşturabilir. Ergenliğe ulaşan erkek filler aile gruplarını terk eder veya yalnız ya da diğer erkek fillerle birlikte bir grup oluşturabilir. Erişkin erkekler çiftleşmek için eş aradıklarında aile grupları ile bir araya gelir. Testosteron salgılanmasının arttığı ve aşırı saldırgan davranışların görüldüğü mest adı verilen durum erkek fillerin baskın olmalarına olanak verir ve üreme başarısını artırır. Aile gruplarında ilgi odağı yavru fillerdir ve anneleri tarafından üç yıl kadar bakılırlar. Filler doğal ortamlarında 70 yıl kadar yaşar; dokunma, görme ve işitme yolu ile iletişim kurar. Uzun mesafelerde, filler insanın duyamayacağı kadar düşük frekanslı sesler ve sismik iletişim yolu ile haberleşir. Fillerin zekâsı primatlar ve balinalar ile kıyaslanacak düzeydedir. Kendilerinin farkında oldukları ve kendi cinslerinden ölmekte olan ya da ölmüş hayvanlara karşı empati gösterdikleri gözlemlenmektedir.
Afrika filleri Dünya Doğa ve Doğal Kaynakları Koruma Birliği (IUCN) tarafından soyunun tükenme riski yüksek olan hassas türler arasında listelenirken, Asya filleri soyunun tükenme riski çok yüksek olan tehlikedeki türler arasında listelenmiştir. Fil popülasyonları için en büyük tehditlerden birisi kaçak olarak avlanan hayvanların dişleri ile yapılan fildişi ticaretidir. Diğer tehditler arasında doğal yaşam alanı kaybı ve yerel halk ile olan çıkar çatışmaları sayılabilir. Filler, Asya'da yük hayvanı olarak kullanılmaktadır. Geçmişte savaşlarda da kullanılan filler günümüzde hayvanat bahçeleri ve sirklerin üyelerindendir. Çok kolay tanınabilen filler sanat, folklor, din, edebiyat ve popüler kültür alanlarında sıklıkla kullanılmıştır.
Adlandırma
Fil adı Türkçeye, Arapçada aynı anlama gelen فيل (fil) kelimesinden geçmiştir. Arapçaya Aramice פיל (pīl), Aramiceye de Sanskritçe "fildişi" anlamına gelen पीलु (pīlu) kelimelerinden geçiş olmuştur.[1] Asya filinin cins adı Elephas Latincede "fil" anlamındadır ve Latinceye, Antik Yunancada "fil" anlamına gelen ἐλέφας (elephas) kelimesinden geçmiştir.[2] Afrika filinin cins adı Loxodonta Antik Yunancada "verev yanlı diş" anlamına gelmektedir.[3]
Taksonomi
Sınıflandırma, türler ve alt türler
.jpg)
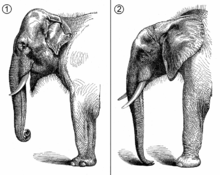
Filler, hortumlular (Proboscidea) takımı içinde yer alan tek familya olan filgiller (Elephantidae) içinde sınıflandırılır. Yaşayan en yakın akrabaları dugonggiller ve manatigillerden oluşan deniz inekleri ve kırsıçanımsıgillerdir. Bu dört familya, Afrotheria üst takımı içinde Paenungulata klâdını oluşturur.[4] Filler ve deniz inekleri ayrıca Tethytheria klâdında gruplandırılırlar.[5] Geleneksel olarak iki fil türü tanınmaktadır: Sahra altı Afrika'da yaşayan Afrika fili (Loxodonta africana) ile Güney ve Güneydoğu Asya'da yaşayan Asya fili (Elephas maximus). Afrika filleri daha büyük kulaklı olur ve sırtları içbükeydir. Ayrıca derileri daha kırışık, karınları eğimli olur ve hortumlarının ucunda iki adet parmak gibi uzantı vardır. Asya fillerinin ise kulakları daha küçük olur. Sırtları dışbükey ya da düzdür. Derileri daha pürüzsüzdür. Karınları yataydır ve ortada bel verir. Hortumlarının ucunda ise tek bir uzantı bulunur. Asya fillerinin azı dişlerindeki çıkıntılar daha dardır ve Afrika fillerinden bu çıkıntılar baklava şeklindedir. Asya fillerinin tepesinde kamburumsu bir çıkıntı bulunur ve derilerinde bölgesel olarak pigment eksikliği görülür.[6]
İsveçli doğabilimci Carl Linnaeus ilk olarak 1758 yılında Elephas cinsini ve o zamanlar Seylan olarak bilinen Sri Lanka'dan bir fili Elephas maximus adıyla, Georges Cuvier 1798 yılında Hint filini Elephas indicus adıyla, Felemenk zoolog Coenraad Jacob Temminck ise 1847 yılında Sumatra filini Elephas sumatranus adıyla tanımladı. İngiliz zoolog Frederick Nutter Chasen ise 1940 yılında bu üç fil türünü de Asya filinin alt türleri olarak sınıflandırdı.[7] Asya fillerinin renkleri ve derilerindeki pigment kaybı coğrafi olarak değişiklik gösterir. Seylan fili (Elephas maximus maximus) Sri Lanka'da yaşar; Hint fili (E. m. indicus) Asya kıtasında Hint altkıtası ve Hindiçin'e özgüdür; Sumatra fili (E. m. sumatranus) ise Sumatra'da bulunur.[6] Tartışmalı olan alt türlerden birisi olan Borneo fili Borneo'nun kuzeyinde yaşar ve diğer alt türlerden daha küçüktür. Kulakları daha büyük ve kuyruğu daha uzundur. Uzun dişleri tipik bir filden daha düzdür. Sri Lankalı zoolog Paules Edward Pieris Deraniyagala bu alt türü 1950 yılında National Geographic dergisindeki bir fil fotoğrafını tip tür alarak Elephas maximus borneensis adıyla tanımlamıştır.[8] Daha sonraları bu alt tür hem E. m. indicus hem de E. m. sumatranus içinde sınıflandırılmıştır. 2003 yılında yapılan genetik analizler sonucunda bu fillerin atalarının anakara popülasyonundan yaklaşık 300.000 yıl önce ayrıldığı gösterilmiştir.[9] 2008 yılında yapılan bir araştırmada ise Borneo fillerinin adaya özgü olmadığı ve günümüzde fillerin soyunun tükendiği Cava Adası'ndan 1521 yılından önce Sulu sultanları tarafından Borneo'ya getirildiği ortaya çıkarılmıştır.[8]

Afrika fili, ilk olarak Alman doğa bilimci Johann Friedrich Blumenbach tarafından 1797 yılında Elephas africana olarak tanımlanmıştır.[10] Loxodonta cinsinin adının Georges Cuvier tarafından 1825 yılında verildiği düşünülmektedir. Cuvier, cinsin adını Loxodonte olarak yazmış ve adı bilinmeyen bir yazar tarafından bu isim Loxodonta olarak düzeltilmiştir. Uluslararası Zoolojik Adlandırma Kodu'nda bu adlandırmanın kaynağı olarak adı bilinmeyen yazar tanınmaktadır.[11] 1942 yılında Henry Fairfield Osborn tarafından Afrika filinin 18 alt türü tanımlanmıştır ancak daha fazla morfolojik veriye ulaşıldıktan sonra alt tür sınıflandırılması azalmıştır[12] ve 1990'larda yalnızca iki alt tür tanınmaya başlanmıştır: savan fili (L. a. africana) ve orman fili (L. a. cyclotis).[13] Orman filinin kulakları daha küçük ve yuvarlaktır, ayrıca uzun dişleri de daha ince ve düzgündür. Yalnızca Batı ve Orta Afrika'nın ormanlık alanlarında bulunur.[14] 2000 yılında yapılan bir inceleme yazısında, kafatası morfolojisine dayanarak bu iki alt türün ayrı birer tür olarak (L. africana ve L. cyclotis) sınıflandırılması önerilmiştir.[15] 2001 ve 2007 yıllarında yayımlanmış olan DNA araştırmaları da iki ayrı tür olarak sınıflandırılması sonucuna varmış[16][17] ancak 2002 ve 2005 yıllarında yapılan DNA araştırmaları bunların aynı tür olduğu sonucuna varmıştır.[18][19] Daha sonra 2010 yılında yapılan bir başka çalışma da Afrika savan ve orman fillerinin ayrı türler olarak sınıflandırılması önerisini desteklemiştir.[20] 2011 yılında Afrika fillerinin taksonomik konumlaması hâlâ tartışmalı bir konudur.[21] Mammal Species of the World (Dünya Memeli Türleri) adlı referans eserin üçüncü baskısında bu iki fil ayrı bir tür olarak sınıflandırılır[11] ve Loxodonta africana türü için alt tür listelemez.[11] Bu yaklaşım Birleşmiş Milletler Çevre Programı'nın Dünya Koruma İzleme Merkezi (UNEP-WCMC) ve Dünya Doğa ve Doğal Kaynakları Koruma Birliği (IUCN) tarafından kabul edilmemiş ve her iki örgüt de L. cyclotis adını L. africana adının sinonimi olarak listelemiştir.[22] Bulunan bazı kanıtlara göre Batı Afrika'da bulunan fillerin ayrı bir tür olduğu önerilmekte olsa da[23] bu hâlâ çözülmemiş bir konudur.[19][21] Kongo Havzası'nın Loxodonta pumilio adı ile ayrı bir tür olması için önerilen pigme filler ise muhtemelen çevresel koşular nedeniyle küçük cüsseli olan ve erken ergenliğe erişen orman filleridir.[24]
.JPG) Seylan fili
Seylan fili
(E. m. maximus) Sumatra fili
Sumatra fili
(E. m. sumatranus) Hint fili
Hint fili
(E. m. indicus) Borneo fili
Borneo fili
(E. m. borneensis) Afrika savan fili
Afrika savan fili
(L. a. africana) Afrika orman fili
Afrika orman fili
(L. a. cyclotis)
Evrimi ve soyu tükenmiş akrabaları


Hortumlular takımının üç ana evrimsel yayılımı ve 161'in üzerinde soyu tükenmiş üyesi kaydedilmiştir. İlk hortumlular, Paleosen'in sonlarında ilk evrimsel yayılımı oluşturan Afrikalı Eritherium ve Phosphatherium cinsleridir.[25] Kalıntıları 1996 yılında Fas'ta bulunan ve yaklaşık tilki büyüklüğünde olan Phosphaterium 56 milyon yıl önce yaşamıştır.[26] Eosen döneminde Hint altkıtasında Anthracobunidae familyası ve Afrika'da Numidotherium, Moeritherium ile Barytherium cinsleri yaşamıştır. Görece küçük olan bu hayvanlar suda yaşıyordu. Moeritherium cinsi hortumlular yaklaşık bir tapir (110 cm) büyüklüğündeydi; uzun burun-dudakları ile kafası domuza benzemekteydi ve ön dişleri uzundu. Fillere benzeyen özellikleri yanında deniz inekleri ile de ortak özelliklere sahipti. Sonraları Phiomia ve Palaeomastodon cinsleri ortaya çıktı. Palaeomastodon muhtemelen ormanlık ve yarı açık ağaçlık alanlarda yaşıyordu. Hortumluların çeşitliliği Oligosen döneminde azalma göstermiştir.[27] Bu dönemin dikkate değer türlerinden biri, daha sonra ortaya çıkacak çeşitli türlerin atası sayılabilecek olan ve Afrika Boynuzu'nda yaşamış olan Eritreum melakeghebrekristosi türüdür.[28] Miyosen'in başlarında Deinotheriidae ve Mammutidae türleri ile birlikte ikinci evrimsel yayılma görüldü. Bunların ilki Afrika ve Avrasya'da yaşayan Barytherium cinsi ile[29] ikincisi de Eritreum cinsi ile akrabadır[28] ve Kuzey Amerika'ya yayılmıştır.[29]
İkinci evrimsel yayılım muhtemelen Eritreum cinsinden türemiş olan[28] Gomphotheriidae familyasının Miyosen döneminde ortaya çıkmasıyla Afrika'da başlamış[29] ve Avustralya ile Antarktika haricinde diğer tüm kıtalara yayılmıştır. Bu grubun üyeleri arasında Gomphotherium ve Platybelodon cinsleri bulunur.[29] Üçüncü evrimsel yayılım ise Miyosen'in sonlarına doğru başlamış ve Gomphotheriidae familyasından türeyen ve uzun bir sürede bunların yerine geçen filimsilerin ortaya çıkışına yol açmıştır.[30] Afrikalı Primelephas gomphotheroides türünden Loxodonta, Mammuthus ve Elephas cinsleri evrimleşmiştir. Bu grubun içinde ilk olarak Loxodonta Miyosen ile Pliyosen arasında diğerlerinden ayrışmış; Mammuthus ile Elephas ise Pliyosen'in başlarında ayrışmıştır. Loxodonta Afrika'da kalırken Elephas Avrasya'ya, Mammuthus ise hem Avrasya'ya hem de Kuzey Amerika'ya yayılmıştır. Aynı zamanda Gomphotheriidae familyasından gelen bir başka hortumlu familyası olan Stegodontidae familyası da Hint alt kıtası, Çin, Güneydoğu Asya ve Japonya olmak üzere Asya'ya yayılmıştır. Mammutidae familyası mastodon gibi yeni türlere evrimleşmiştir.[31]

Buzul Çağı'nın başında filimsiler yüksek oranda türleşme sürecine girmiştir. Loxodonta atlantica Afrika'nın kuzeyi ve güneyinde en yaygın filimsi türü olmuş ancak Buzul Çağının sonlarına doğru yerine Elephas iolensis türü geçmiştir. Elephas cinsi Afrika'dan tamamen yok olduktan sonra Loxodonta cinsi çağdaş türlerle baskın hâle gelmiştir. Elephas cinsi Asya'da E. hysudricus ve çağdaş Asya filinin atası sayılabilen E. platycephus[32] gibi yeni türlere evrimleşmiştir.[33] Mammuthus cinsi aralarında çok tanınmış olan tüylü mamut da dahil olmak üzere çeşitli türlere ayrılmıştır.[33] Buzul Çağının son döneminde hortumlu türlerinin çoğu Dünya üzerinde 5 kg'ın üzerinde olan cinslerin %50'sinin yok olduğu buzulların ilerlemesi sırasında ortadan kaybolmuştur.[34]
Hortumlular çeşitli evrimsel eğilimlere maruz kalmıştır. Cüsselerinin büyümesi bunlardan biridir ve bu nedenle boyları 4 m'yi aşan dev türler ortaya çıkmıştır.[35] Aralarında soyu tükenmiş olan Sauropoda alt takımından dinozorların da bulunduğu diğer megaotçullar gibi fillerin büyük cüsseye sahip olmaları, düşük besin değeri olan bitkisel besinlerle yaşayabilmeleri içindir.[36] Uzuvları uzadı ve ayakları daha kısaldı ve kalınlaştı. İlk hortumluların alt çeneleri daha uzun ve kafatasları daha küçük iken daha gelişmiş olan hortumluların alt çeneleri daha kısadır ve böylece kafalarının kütle merkezi değişmiştir. Kafatası büyümüş ve boyun kafatasına daha iyi destek sağlamak için kısalmıştır. Cüssenin artması sonucunda hareketli olan hortum da daha uzağa erişebilmek için uzamıştır. Ön azı, kesici ve köpek dişlerinin sayısı azalmıştır. Azı dişleri daha büyümüş ve daha özelleşmiştir. Üst ikinci kesici dişler uzamış ve düz, yukarı ya da aşağı eğimli ya da spiral şeklinde olmak üzere türlere göre farklılık göstermiştir. Bazı hortumluların üst yerine alt kesici dişleri uzamıştır.[35] Filler, orta kulak anatomileri ve testislerinin içeride olması gibi suda yaşayan atalarından gelen bazı özellikleri korumuşlardır.[37]
Mammuthus cinsinin Loxodonta ya da Elephas cinsleri ile olan yakınlığı konusunda bazı tartışmalar olmuştur. Bazı DNA araştırmaları Mammuthus cinsinin Loxodonta cinsine daha yakın olduğunu[38][39] diğerleri ise Elephas cinsine yakın olduğunu[5] gösterir. Ancak tüylü mamutun mitokondriyal DNA profilinin tamamının 2005 yılında yapılan analizi Mammuthus cinsinin Elephas cinsi ile daha yakın akraba olduğunu kanıtlar.[16][20][40] Morfolojik kanıtlar Mammuthus ve Elephas cinslerini yakın akraba olarak gösterirken albümin proteinleri ve kollajen karşılaştırması her üç cinsin de birbirine aynı oranda yakın olduğuna işaret eder.[41] Bazı bilim insanları bir gün klonlama yolu ile yaratılacak mamut embriyosunun Asya filinin rahmine yerleştirilebileceğine inanmaktadırlar.[42]
Cüce türler

Adalarda yaşayan çeşitli hortumlu türler zamanla ada cüceleşmesine maruz kalmıştır. Her ne kadar Pliyosen'den önce cüce fil türleri var olmuşsa da bu süreç Buzul Çağı sırasında değişen deniz seviyeleri nedeniyle bazı fil popülasyonlarının adalarda izole olmasıyla gerçekleşmiştir. Bu fillerin cüsseleri, adalarda büyük yırtıcıların olmaması ve sınırlı kaynaklar bulunması nedeniyle küçülmüştür. Benzer koşullar altında kemiriciler gibi küçük memeliler ise ada devleşmesi sürecine maruz kalır. Cüce hortumlu türlerin Endonezya'da, Kaliforniya Channel Adaları'nda ve Akdeniz'in çeşitli adalarında yaşadıkları bilinmektedir.[43]
Sulawesi'de yaşamış olan Elephas celebensis türünün Elephas planifrons türünden geldiği düşünülmektedir. Malta ve Sicilya'da 'yaşamış olan Elephas falconeri türünün boyu yalnızca 1 m civarındaydı ve muhtemelen Elephas antiquus türünden evrimleşmişti. Elephas antiquus türünden gelen diğer cüce fillere Kıbrıs'ta rastlanır. Girit, Kiklad Adaları ve Oniki Ada'da yaşamış olan cüce fillerin hangi türden evrimleştiği bilinmemektedir.[43] Sardinya'da cüce mamutların yaşadığı bilinmektedir.[43] Mammuthus columbi Kaliforniya Channel Adaları'na yerleşmiş ve cüce mamut türü olan Mammuthus exilis türüne evrimleşmiştir. Bu türün boyu 1,2 ilâ 1,8 m civarında ve ağırlığa da 200 ilâ 2.000 kg arasındaydı. Vrangel Adası'nda 4.000 yıl kadar önceye kadar, küçük cüsseli tüylü mamut popülasyonunun yaşamakta olduğu ortaya çıkarılmıştır.[43] 1993'te kalıntıları bulunduktan sonra cüce mamut olarak sınıflandırıldılar.[44] Bu sınıflandırma daha sonra tekrar gözden geçirilmiş ve 1999'dan beri gerçek "cüce mamut" olarak kabul edilmemişlerdir.[45]
Anatomi ve morfoloji
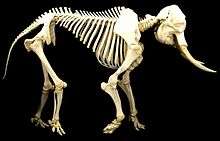
Fil karada yaşayan en büyük hayvandır. Afrika fillerinin boyları 3 ilâ 4 m arasında, ağırlıkları da 4.000 ilâ 7.000 kg arasında; Asya fillerinin boyu ise 2 ilâ 3,5 m ve ağırlıkları da 3.000 ilâ 5.000 kg arasında değişir.[6] Her iki cinsin erkekleri dişilerinden daha büyüktür.[7][10] Afrika filleri arasında orman ırkı, savan ırkından daha küçüktür.[14] Filin iskeleti 326 ilâ 351 kemikten oluşur.[46] Omurların birbirine sıkı eklemlerle bağlı olması nedeniyle sırtın esnekliği sınırlıdır. Afrika fillerinin 21 çift kaburgasına karşın Asya filleri 19 ilâ 20 çift kaburgaya sahiptir.[47]
Fil kafatası, fildişlerinden kaldıraç etkisiyle gelen kuvvetlerle kafa kafaya çarpışmalarda oluşan kuvvetlere dayanabilecek kadar esnektir. Kafatasının arkası düzleşmiş ve yayılmış ve beyni her yönden koruyabilecek şekilde kemerli bir yapıya kavuşmuştur.[48] Kafatasında bulunan sinüsler (hava boşlukları) sayesinde kafatasının ağırlığı azalırken toplam mukavemeti korunmaktadır. Bu boşluklar, kafatasının içinin petekler gibi görünmesine yol açar. Oldukça geniş olan kafatası, tüm kafanın taşınabilmesi için kasların tutunabileceği bir yüzey sağlar. Alt çene ağır ve sağlamdır.[46] Kafanın boyutları yüzünden boyun görece kısadır ve iyi bir destek sağlar.[35] Filin gözünde gözyaşı sistemi olmadığı için gözün nemli kalması Harder bezi sayesinde olur. Dayanıklı üçüncü göz kapağı, göz küresini korur. Gözlerin kafa üzerindeki konumu ve hareket sınırlılığı hayvanın görüş alanını kısıtlar.[49] Filler dikromattır[50] ve loş ışık altında iyi görmelerine rağmen parlak ışık altında çok iyi görmezler.[51] Vücut sıcaklığı, insana benzer ve ortalama 35,9 °C 'dir. Fil de deve gibi aşırı çevresel koşullara uyum sağlayabilmek için vücut ısısını birkaç derece artırıp azaltabilir.[52]
Kulaklar

Fil kulaklarının tabanı kalın uçları incedir. Kulak kepçesi bolca kılcal damar içerir. Kulak kepçesindeki kılcal damarlardan akan ılık kan, çevreye ısısını vererek vücut sıcaklığının düşmesini sağlar. Bu durum kulak kepçesi hareketsiz dururken oluşur ve fil kulaklarını hareket ettirerek sıcaklığın düşmesinin daha hızlı olmasını sağlayabilir. Daha büyük kulak yüzeyi daha çok ısının çıkabilmesine olanak verir. Tüm fil türleri içinde Afrika savan filleri en sıcak iklimde yaşar ve en büyük kulak kepçelerine sahiptir.[53] Filler düşük frekanslarda duyabilirler ve en hassas oldukları aralık 1 kHz civarındadır.[54]
Hortum
Filin hortumu burun ile üst dudağın kaynaşmasından oluşur. Fil fetusunun hortumu ve üst dudağı ayrıktır.[35] Uzun ve uzmanlaşmış olan hortum filin en önemli ve çok yönlü uzvudur. İçinde çok az yağ bulunan ve hiç kemik bulunmayan hortumda 150.000 kadar ayrı kas demeti bulunur. Bu eşleşmiş kaslar, yüzeysel ve iç kaslar olarak iki çeşittir. Yüzeysel kaslar dorsal, ventral ve lateral olarak üçe ayrılır. İç kaslar da çapraz ve ışınsal olarak ikiye ayrılır. Hortum kasları, kafatasında bulunan kemiksi bir açıklığa bağlıdır. Burun bölmesi burun delikleri arasında yatay olarak uzanan küçük kaslardan oluşur. Burun bölmesi tabanı kıkırdak ile ikiye ayrılır.[55] Kassal hidrostat olan hortum hassas olarak koordine olan kas kasılmalarıyla hareket eder. Kasların hareketleri hem birlikte hem de karşılıklı olarak gerçekleşebilmektedir. Maksiler ve fasiyal sinirin oluşturduğu tek bir hortum siniri hortumun iki yanı boyunca uzanır.[56]
Fil hortumu soluma, koku alma, dokunma, tutma ve ses çıkarma gibi çeşitli işlevlere sahiptir.[35] Filin koku alma duyusu tazınınkinden dört kat daha güçlüdür.[57] Hortumun güçlü sarma ve bükme yapabilme yeteneği besin toplamasına, diğer fillerle güreşebilmesine[58] ve 350 kg'a kadar ağırlık kaldırabilmesine olanak tanır.[35] Aynı zamanda göz silme ya da bir deliği kontrol etme gibi hassas görevlerde de kullanılabilir. Bir fil hortumuyla yer fıstığına zarar vermeden kabuğunu kırabilir,[35] 7 m'ye ulaşan yüksekliklerdeki nesnelere erişebilir ve çamur ile kum altında su arayabilir.[58] Hortumla nesneleri kavramada bireyler arasında farklı yön tercihleri görülür, bazıları sağa bazıları da sola doğru hortumlarını kıvırmayı tercih eder.[56] Fil, hortumuyla içmek ya da vücuduna püskürtmek için su çekebilir.[35] Yetişkin bir Asya fili hortumu ile 8,5 litreye kadar su tutabilir.[55] Suyun dışında vücutlarına toz ve ot da püskürtürler.[35] Suyun altındayken hortumunu şnorkele benzer şekilde kullanıp, nefes alıp verebilirler.[37]
Afrika filinin hortumunun ucunda, besinleri tutup ağzına götürmesini sağlayan ve parmağa benzeyen iki adet uzantı bulunur. Asya filinde bu uzantı bir tanedir ve besinleri daha çok hortumun ucu ile sarıp sıkıştırarak ağzına götürür.[6] Asya fillerinin kas koordinasyonu Afrika fillerine oranla daha iyidir ve karmaşık görevleri yerine getirebilirler.[55] Filin hortumunun kesilmesi hayatta kalmasını zorlaştırırsa da[35] nadir durumlarda kısalmış hortum ile yaşamlarını sürdürebilmiş örnekler bulunmaktadır. Böyle bir filin ön ayakları üzerinde diz çökerek ve arka ayaklarını kaldırarak dudakları yardımıyla otladığı gözlemlenmiştir.[55] Gevşek hortum sendromu Afrika savan fillerinde görülen ve uçlardan başlayarak çevresel sinirler ile kasların bozulmasıyla oluşan bir hortum felci durumudur.[59]
 Hortumunu kaldırmış olan Afrika fili.Fil hortumuyla ses çıkarırken genellikle yukarıya kaldırır
Hortumunu kaldırmış olan Afrika fili.Fil hortumuyla ses çıkarırken genellikle yukarıya kaldırır Asya fili'nin (A) hortumunun ucunda tek parmak gibi uzantı bulunurken Afrika filinin (B) hortumunun ucunda iki uzantı bulunur.
Asya fili'nin (A) hortumunun ucunda tek parmak gibi uzantı bulunurken Afrika filinin (B) hortumunun ucunda iki uzantı bulunur. Fil hortumu ile gözünü temizleyecek kadar hassas ve nazik davranabilir.
Fil hortumu ile gözünü temizleyecek kadar hassas ve nazik davranabilir. Fil hortumu ile 8,5 litre su tutabilir.
Fil hortumu ile 8,5 litre su tutabilir. İnsan elinden hortumuyla beslenen dişi Asya filinin hortumunun ucu.
İnsan elinden hortumuyla beslenen dişi Asya filinin hortumunun ucu.
Dişler
.jpg)
Uzun olan ve fildişi olarak bilinen kesici dişler, 12 önazı sütdişi ve 12 azı dişi olmak üzere fillerin genellikle 26 dişi vardır. Bebek dişleri çıktıktan sonra yerine kalıcı dişler gelen memelilerin aksine fillerin yaşamı boyunca dişleri belirli bir döngü ile değişir. Filin tipik yaşam süresi boyunca çiğneme dişleri altı kere değişir. Dişler, memelilerin çoğunda olduğu gibi çeneden dikey olarak çıkıp eski dişlerin yerine geçmez. Bunun yerine yeni dişler ağzın arkasında çıkar ve eski dişleri ağzın önüne doğru iter. Çenenin her iki yanındaki ilk çiğneme dişleri fil iki ya da üç yaşına geldiğinde, ikinci dişler fil dört ilâ altı yaşındayken düşer. Üçüncü dişler 9-15 yaşında, dördüncü dişler ise 18-28 yaşında değiştirilir. Beşinci dişler fil 40'lı yaşlarının başındayken düşer. Altıncı ve genellikle de sonuncu dişler ile fil yaşamının sonuna kadar yaşar. Fil dişlerinin tepeleri halka şeklinde olup Afrika fillerinde bu tepeler baklava şeklindedir.[60]
Fildişleri

Fildişleri üst çenede bulunan kesici dişlerin değişiklik geçirmiş hâlidir. Hayvan 6 ilâ 12 aylıkken sütdişlerinin yerine geçer ve yılda yaklaşık 17 cm olmak üzere uzamaya devam eder. Yeni çıkan fildişlerinin üzerinde bulunan pürüzsüz mine tabakası zamanla aşınır. Dentin fildişi olarak bilinir ve kesiti çapraz çizgilerden baklava şeklinde alanlar oluşturan bir desendir. Yaşayan bir doku olarak fildişi görece yumuşaktır ancak kalsit kadar sertliğe sahiptir. Kesici dişlerin büyük kısmı dışarıdan görünür, kalan kısmı kafatasındaki oyuklara bağlıdır. Kesici dişlerin en azından üçte birinde pulpa bulunur ve bazılarında uçlara kadar giden sinirler de görülür. Bu nedenle hayvana zarar vermeden dişleri sökmek zordur. Dişler söküldükten sonra eğer serin ve nemli yerde tutulmazsa kurumaya ve çatlamaya başlar. Fil, fildişlerini çeşitli amaçlar için kullanır. Su, tuz ve köklere ulaşmak için toprağı kazmaya; ağaçların kabuğunu soyup iz bırakmaya ve yol açmak için ağaçlar ile dalları kaldırmaya yarar. Dövüşürken saldırmak, savunmak ve hortumu korumak için kullanılır.[61]
İnsanların el kullanımında sağ ya da solu tercih etmesi gibi filler de fildişlerini kullanırken bireysel olarak sol ya da sağı tercih ederler. Baskın olan kesici diş genellikle daha çok yıpranmıştır, diğerine göre daha kısadır ve ucu daha yuvarlaktır. Afrika fillerinde hem erkeklerde hem de dişilerde uzun kesici dişler bulunur ve 3 m uzunluğa erişir.[61] Erkeklerin fildişleri dişilere göre daha kalındır.[62] Asya türlerinde yalnızca erkeklerin uzun kesici dişleri vardır. Dişilerin kesici dişleri ya küçüktür ya da hiç yoktur.[61] Uzun kesici dişleri olmayan erkekler de görülür, özellikle Seylan filleri arasında yaygındır.[63] Asya fili erkeklerinin dişleri Afrika fili kadar uzun olabilir ancak daha ince ve hafiftir. Asya filleri arasında ölçülmüş en uzun kesici diş 3,02 m boyunda ve 39 kg ağırlığındadır. Afrika'da [64] ve Asya'da[65] fillerin dişleri nedeniyle avlanması sonucu oluşan doğal seçilim ile fillerde daha kısa dişler baskınlaşmıştır.[66]
Deri

Filin derisi çok dayanıklıdır ve sırt ile başın bazı bölgelerinde 2,5 cm kalınlığa erişir. Ağzın, anüsün ve kulakların içindeki deri kısmen daha incedir. Fillerin derileri genel olarak gri olsa da, Afrika filleri renkli çamurda yuvarlandıktan sonra kahverengi ya da kızılımsı görünebilirler. Asya fillerinin derilerinde kısmen pigment olmayan bölgeler bulunur. Özellikle alın ve kulak çevresinde bu bölgeler görülür. Yavru fillerin kılları, özellikle kafa ve sırt bölgesinde kahverengimsi ya da kızılımsı olur. Filler olgunlaştıkça kılları koyulaşır ve azalır; ancak kuyruğun ucunda, çenede, cinsel organlar ile göz ve kulak çevresinde yoğun kıl öbekleri bulunur. Normal olarak Asya filleri Afrika fillerinden daha kıllıdır.[67]
Filler morötesi ışıktan kendilerini korumak için çamurdan yararlanırlar. Kalın olmasına rağmen derileri çok hassastır. Güneş yanığından, böcek ısırığından ve nem kaybından kendilerini korumak için düzenli çamur banyosu yapmazlarsa derileri ciddi zarar görebilir. Fil, suda yıkandıktan sonra hortumuyla gövdesini toz ile kaplar ve bu toz tabakası kuruyarak koruyucu bir tabaka oluşturur. Fillerin hacimlerine göre derilerinin yüzey alanı çok düşük olduğundan derilerinden ısı kaybında çok zorlanırlar. Tabanlarının hava ile temas etmesi için bacaklarını kaldırdıkları gözlemlenmiştir.[67]
Bacaklar, hareket ve duruş

Filin ağırlığını taşıyabilmek için bacakları diğer memelilere göre gövdesinin altında daha dik olarak konumlanmıştır. Uzun bacak kemiklerinde ilik boşluğu yerine süngerimsi kemik yapısı bulunur. Bu yapı kan oluşumuna olanak verirken kemiklerin daha dayanıklı olmasını sağlar.[68] Ön ya da arka ayaklar hayvanın ağırlığını taşıyabilir ama genellikle ağırlığın %60'ı ön ayaklar tarafından taşınır.[69] Bacak kemikleri birbirlerinin üzerinde ve doğrudan gövdenin altında olduğu için fil çok enerji harcamadan uzunca süre hareketsiz durabilir. Ulna ve radius pronasyon konumunda bağlı oldukları için filler ön ayaklarını döndüremezler; manusun ayası geriye doğru bakar.[68] Pronator quadratus ve pronator teres kasları ya çok küçülmüştür ya da yok olmuştur.[70] Fillerin yuvarlak ayaklarında manus ve pes altında hayvanın ağırlığının dağıtılmasına yarayan yumuşak doku bulunur.[69] Dev pandanın fazladan olan "başparmağı"na benzer susamsı kemikten ibaret ve ağırlık dağılımına yardımcı olan fazladan bir parmakları vardır.[71] Hem arka hem de ön ayaklarda beş adet tırnak vardır.[6]
Filler hem ileriye hem de geriye doğru hareket edebilirler ancak zıplayamaz, tırıs ya da dörtnala gidemezler. Hareket ederken biri yürüme diğeri de koşma sayılabilecek iki hız kullanabilirler.[68] Yürürken bacaklar sarkaç gibi hareket eder, ayak yere konulurken omuzlar ve kalçalar yükselip alçalır. Hızlı yürümede filler bacaklarını diğer koşan hayvanlar gibi kullansa da ayakların yerden kesildiği bir anı olmadığı için hızlı yürüme tam olarak koşma kıstaslarını sağlamaz.[72] Hızlı hareket eden filler ön ayaklarıyla koşuyor, arka ayaklarıyla yürüyor gibi görünürler ve yaklaşık saatte 18 km'lik bir hıza ulaşabilirler.[73] Bu hızlarda diğer dört ayaklıların çoğu dörtnala hareket eder. Yay kinetiği fillerin hareketinin diğer hayvanlardan farkını açıklayabilir.[74] Hareket ederken filin ayağı altındaki yumuşak doku genişleyip daralır ve bu kadar ağır bir hayvanın hareket etmesinden kaynaklanacak olan sesi ve ağrıyı azaltır.[69] Filler iyi yüzücüdür. Dibe değmeden altı saat kadar yüzebildikleri, bir kerede 48 km mesafe katedebildikleri ve 2,1 km/s hız yapabildikleri kaydedilmiştir.[75]
İç ve cinsel organlar
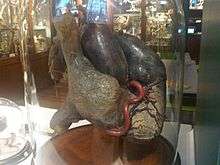
Filin beyni 4,5 ilâ 5,5 kg ağırlığındadır. Ağırlığı 1,6 kg olan insan beyni ile karşılaştırıldığında filin beyni daha büyük olsa da oransal olarak daha küçüktür. Doğduktan hemen sonra fil beyni erişkinlikte ulaşılan ağırlığın %30 ilâ 40'ı arasındadır. Serebrum ve beyincik iyi gelişmiştir ve temporal loblar o kadar büyüktür ki yanlara doğru kabarıklık yapar.[52] Filin boğazında daha sonra kullanmak üzere su saklayabileceği bir kese bulunur.[35]
Filin kalbi 12 ilâ 21 kg arasındadır. Memeliler arasında sıradışı bir şekilde kalp apeksi çift uçludur.[52] Ayakta iken filin kalbi dakikada 30 kere atar. Diğer hayvanların tersine fil yattığında kalp atışı dakikada 8 ilâ 10 kere olmak üzere artış gösterir.[76] Akciğerler diyaframa bağlıdır[52] ve akciğer zarı yerine bağ doku bulunur.[37] Bu nedenle soluma göğüs kafesinin genişlemesinden çok diyaframa bağlıdır.[52] Bu hayvanın su altındayken hortumuyla yüzeyden nefes alması sırasında oluşan basınç farklılıklarıyla başedebilmesine olanak sağlar[37] ancak bu açıklamanın geçerliliği tartışmalıdır.[77] Bu adaptasyonun bir başka açıklaması da hayvanın hortumu ile su çekebilmesine yardımcı olmasıdır.[37] Filler genellikle hortumlarıyla soluk alır ancak çok az bir hava ağızlarından da geçer. Geviş getirmeyen ve tek bölmeli mideye sahip olan fillerde arka bağırsak mayalanması ile selüloz çekum ve kalın bağırsakta sindirilir. İnce, kör ve kalın bağırsaklarının toplam uzunluğu 35 m'ye yaklaşır. Sindirim bir gün kadar sürse de fillerin ağızdan aldıkları besinlerin çoğu sindirilmeden dışkılanır.[52]
Erkek filin testisleri gövdenin içinde böbreklerin yanındadır. Fil penisinin boyu 100 cm'ye, kalınlığı da tabanında 16 cm'ye ulaşır. Ereksiyonda iken S şeklindedir ve ucunda Y şeklinde yarık vardır. Dişi filin gelişmiş klitorisi 40 cm uzunluğundadır. Vulva çoğu memelide olduğu gibi kuyruğa yakın olmak yerine arka ayakların arasındadır.Hayvanın büyük karın boşluğu nedeniyle gebeliği tespit etmek kolay değildir. Dişinin meme bezleri ön ayaklarının arasındadır, dolayısıyla süt emen yavru dişinin hortumunun ulaşabileceği mesafededir.[52] Fillerde başın iki yanında yer alan kendine has temporin salgılayan bir bez vardır. Bu organ cinsel davranış ile bağlantılıdır ve erkekler kızgınlığa girdiklerinde buradan bir sıvı salgılar.[78] Dişilerin de aynı yerden salgı yaptıkları gözlemlenmiştir.[57]
Davranış ve yaşam
Ekoloji

Afrika savan fili savanlar, çöller, bataklıklar ve göl kıyıları gibi çeşitli doğal alanlarda ve deniz seviyesinden kar hattının üzerinde dağlara kadar her türlü rakımda yaşar. Orman filleri ise ekvator ormanlarında yaşar ama galeri ormanları ile ormanlar ve savanlar arasındaki ekotonlara da girebilirler.[14] Asya filleri otlak, fundalık ve ağaçlardan oluşan alanları tercih eder, Hindistan'ın güneyi ve Sri Lanka'da dikenli fundalık ormanlarda, Malay Yarımadası'nda da herdem yeşil yapraklı ormanlarda yaşarlar.[7] Filler otçuldur ve yapraklar, sürgünler, meyveler, ağaç kabukları ve kökleri ile beslenirler. Afrika filleri daha çok yaprak, sürgün ve meyvelerle beslenmeyi tercih ederken Asya filleri yere yakın otlar ve köklerle beslenmeyi tercih eder. Günde yaklaşık 150 kg kadar besin tüketir ve 40 litre kadar su içerler. Filler genellikle su kaynaklarına yakın kalmayı tercih eder.[14] Asıl olarak sabah, öğleden sonra ve gece beslenirler. Öğle vakti filler ağaçlar altında dinlenir ve ayakta kestirebilirler. Asıl geceleri yere yatarak uyurlar.[68][79] Günde ortalama 3 ilâ 4 saat arası uyurlar.[80] Hem erkekler hem de aile grupları tipik olarak günde 10 ilâ 120 km arasında yol kat edebilir ancak Namibya'da Etosha Ulusal Parkı'nda 90 ilâ 180 km kadar uzun mesafe katettikleri de kaydedilmiştir.[81] Besin, su ve eş aramak için mevsimsel olarak göç edebilirler. Botsvana'da Chobe Ulusal Parkı'nda fil sürülerinin yerel su kaynakları kuruduğunda 325 km yol alarak nehre indikleri gözlemlenmiştir.[82]

Büyük cüsselerinden ötürü filler çevreleri üzerinde çok önemli etkide bulunur ve bu nedenle kilittaşı türlerden sayılır. Ağaçları ve çalılıkları köklerinden sökme âdetleri savanları otlaklara çevirebilir. Kuraklık sırasında su bulmak için kazdıkları çukurlardan diğer hayvanlar da yararlanır. Açtıkları su çukurlarını genişleterek çamur içinde yuvarlanırlar. Elgon Dağı'nda fillerin kazdığı mağaralar toynaklılar, kırsıçanımsılar, yarasalar ve böcekler tarafından kullanılır.[83] Filler önemli tohum dağıtıcısıdırlar. Afrika orman filleri tohumları yutar ve dışkı ile dışarı çıkarır. Bu sürecin çimlenmeye etkisi ya yoktur ya da olumlu etkisi vardır. Tohumlar büyük mesafelerde dağıtılır.[84] Asya ormanlarında büyük tohumlar dağılmak için filler ve gergedanlar gibi dev otçullara gereksinim duyar. Bu ekolojik niş bir sonraki en büyük otçul olan Hint tapiri tarafından doldurulamaz.[85] Fillerin yedikleri besinlerin çoğu sindirilmeden atıldığı için fil dışkısı bok böceği ve maymunlar gibi diğer hayvanlar için besin kaynağı oluşturur.[83] Ekosistemler üzerinde fillerin olumsuz etkileri de bulunur. Uganda'da Murchison Falls Ulusal Parkı'nda fil popülasyonunun fazlalığı ormanlık araziye bağımlı olan küçük kuş popülasyonlarını tehdit altına sokmuştur. Ağırlıkları nedeniyle toprak sıkışarak yağmurun toprak içine süzülmeden üzerinden akmasına neden olarak erozyonun oluşmasında önemli bir etken olur.[79]

Filler genellikle diğer otçullarla birlikte sakin bir şekilde yaşar ve diğer hayvanlar fillerden uzak durur. Filler ile gergedanlar arasında bazı saldırgan etkileşimler kaydedilmiştir. Kenya'da Aberdare Ulusal Parkı'nda yavru bir file saldıran gergedan aile grubundaki diğer filler tarafından öldürülmüştür.[79] 1990'larda Güney Afrika'da Hluhluwe-iMfolozi Parkı'na getirilen genç öksüz filler rastgele gergedanları öldürmeye başlamış ve daha yaşlı erkek fillerin getirilmesine kadar 36 gergedanı öldürmüştür.[86] Erişkin filler cüsseleri sayesinde diğer yırtıcılara karşı hemen hemen tamamen yenilmez sayılabilirler. Yavru filler Afrika'da aslanlara, benekli sırtlanlara ve yaban köpeklerine[10] Asya'da ise kaplanlara[7] yem olabilmektedir. Botsvana'da Chobe Ulusal Parkı'nda kuraklık döneminde aslanlar filleri avlamayı âdet hâline getirmiştir ve 30 aslandan oluşan bir sürünün dört ilâ onbir yaşlarındaki filleri öldürdüğü kaydedilmiştir.[87] Fillerde, özellikle yuvarlak solucanlar olmak üzere diğer otçullara nazaran çok sayıda parazit görülür.[88]
Sosyal yapı

Erkek ve dişi fillerin sosyal yaşamları birbirinden çok farklıdır. Dişiler yaşamlarını sıkı bağlarla bağlı oldukları anaerkil aile gruplarında geçirir. Bu aile grupları anneler ve yavrularından oluşur ve bazen on bireyden fazla filden oluşur. En yaşlı dişi gruba liderlik eder.[89] Grubun lideri olan dişi ölene kadar[10] ya da bu rolü üstlenmek için enerjisi kalmayana kadar[90] grubun başında kalır. Hayvanat bahçesinde yaşayan filler arasında yapılan bir araştırmada grubun lideri olan dişi öldüğünde diğer fillerin dışkılarında kortikosteron (stres hormonu) düzeyinin dikkat çekici şekilde arttığı gözlemlenmiştir.[91] En yaşlı dişi öldüğünde ya da liderliği bırakmak zorunda kaldığında, kardeşi grupta olsa bile yerine en yaşlı kızı geçer.[10] Yaşlı dişi liderlerin daha etkili karar verici olduğu görülmüştür.[92]
Dişi filin sosyal çevresi yalnızca küçük aile grubu ile sınırlı değildir. Kenya'da Amboseli Ulusal Parkı'nda yaşayan fillere bakıldığında bir dişi filin yaşamı boyunca diğer aile grupları, klanlar ve alt popülasyonlar ile etkileşime girdiği gözlemlenmiştir. Aileler birbirleriyle bağlantıya girip genellikle iki aile grubundan oluşan bağlı gruplar oluşturur. Kurak mevsimde fil aileleri bir araya gelerek klan adı verilen başka bir sosyal yapı oluşturur. Bu klanlar içindeki gruplar sıkı ilişkiler kurmazlar ancak bölgelerini kurak mevsimde diğer klanlara karşı korurlar. Tipik olarak bir klan içinde dokuz grup bulunur. Amboseli fil popülasyonu ayrıca "merkezî" ve "çevresel" olarak iki alt popülasyona da ayrılır.[89]

Hindistan ve Sri Lanka'da yaşayan bazı fil popülasyonlarının da benzer sosyal yapıları vardır. Sıkı aile birimleri ve çok bağlı olunmayan daha büyük topluluklar olduğu görülür. Bebek fillere bakıcılık ve genç fillere koruyuculuk yapılan birimler olduğu gözlemlenmiştir. Hindistan'ın güneyinde fil popülasyonları içinde aile grupları, bağlı aile grupları ve muhtemelen klan yapıları olabilir. Aile grupları bir ya da iki dişi ve yavrularından oluşan küçük gruplardır. İki dişi ve yavrularından daha fazla sayıda bireyi içeren gruplar ortak aile grubu olarak bilinir. Malay fil popülasyonlarının aile birimleri daha küçüktür ve aile ile bağlı aile gruplarından daha büyük sosyal yapı içermezler. Afrika orman fillerinin aile grupları ise genellikle bir dişi ve iki ilâ üç yavrusundan oluşur. Özellikle orman içinde yol açma gibi konularda bu aile gruplarının yardımlaştıkları görülür.[89]
Erişkin erkeğin yaşamı ise dişilerden çok farklıdır. Olgunlaştıkça erkek fil grubundan uzaklaşarak başka erkeklerle ve hatta başka aile gruplarıyla etkileşime geçebilir. Kenya'da Amboseli'de genç erkekler 14 ilâ 15 yaşlarına doğru zamanlarının %80'ini ailelerinden uzakta geçirir. Aile grubunun erişkin dişileri erkek file karşı saldırganca davranışlar sergiler ve bu da erkekleri tamamen gruptan ayrılmaya sevk eder. Erkekler gruptan ayrılınca ya yalnız ya da diğer erkeklerle birlikte yaşar. Genellikle yoğun ormanlarda bulunan erkekler yalnız yaşamayı tercih eder. Asya fillerinin erkekleri genellikle yalnız yaşar ama iki ilâ daha çok erkek filden oluşan gruplar hâlinde de yaşayabilirler. Böyle gruplar arasında en çok yedi erkek filin olduğu bir grup gözlemlenmiştir. İçinde 10 erkek filden fazla üye bulunduran gruplar Afrika savan fillerinde görülür. Bunların en büyüğü içinde 144 Afrika savan fili barındırmaktaydı.[93] Tek ya da grup hâlinde yaşasalar bile erkekler arasında bir baskınlık hiyerarşisi bulunur. Baskınlık yaşa, cüsseye ve cinsel duruma bağlıdır.[93] Yaşlı erkek filler genç erkek fillerin saldırgan davranışlarını kontrol altında tutar ve genç erkeklerin "çete"ler kurmasını engeller.[94] Erişkin erkek ve dişi fil üremek için bir araya gelir. Erkekler içinde östrus hâlinde olan bir dişi bulunan gruplar ile etkileşime girebilir.[93]
Cinsel davranış
Mest

Erişkin erkek filler testosteron hormonunun arttığı mest olarak bilinen bir duruma girer. Güney Hindistan'da bulunan fil popülasyonunda erkek fillerin ilk olarak 15 yaşında mest durumuna girdiğini ancak 25 yaşından önce bu durumun görece daha az etkili olduğu gözlemlenmiştir. Kenya'da Amboseli parkında 24 yaşın altındaki erkek fillerin mest hâline girmediği ve 25 ilâ 35 yaş arasındakilerin yarısı ile 35 yaş üstündekilerin tamamının mest olduğu gözlemlenmiştir. Genç erkeklerin kurak mevsimde (Ocak ile Mayıs arası) yaşlı erkeklerin de yağmur mevsiminde (Haziran ile Aralık arası) mest durumuna girdikleri görülür. Erkek filin mest olduğunun ana göstergesi temporin bezinden salgılanan sıvının yüzlerinin iki yanına doğru akmasıdır. Penisi prepüsün içindekeyken idrarını yapabilir ve bu durumda idrarı arka bacaklarının üzerine sıçrar. Mest durumu ile bağlı davranışlar kafayı yukarda ve sağa sola sallayarak yürüme, fildişleriyle yeri kazma, işaretleme, hortum ile gürleme ve yalnızca tek kulağı sallama gibi davranışlardır. Mest durumu bir gün ile dört ay arasında sürebilir.[95]
Mest durumunda erkek filler aşırı derecede saldırganlaşır. Mest durumunda olan ya da olmayan erkekler arasında agonistik karşılaşmalarda cüsse belirleyici etkendir. İki gruptan bireyler arasındaki karşılaşmaları, mest olmayan erkek daha iri de olsa, çoğunlukla mest durumunda olan birey kazanır. Erkek fil kendinden daha büyük mest durumunda bir fil ile karşılaştığında kendi mest davranışlarını durdurabilir. Benzer büyüklükte olanlar birbirleriyle karşılaşmaktan kaçınır. Agonistik davranışlar genellikle tehdit gösterisinde bulunma, kovalama ve fildişleri ile hafif çarpışmalardır. Ciddi dövüşler nadir görülür.[95]
Çiftleşme

Filler polijinik olarak ürer[96] ve çiftleşmeler yağmur mevsiminin ortasında sıklaşır.[97] Östrusta olan dişi fil idrarı ve vajinal salgılarıyla feromon adı verilen kimyasal sinyaller göndererek çiftleşmeye hazır olduğunu bildirir. Erkek fil potansiyel bir eşi izler ve durumunu flehmen tepkisi ile değerlendirir. Fillerde flehmen tepkisi erkeğin hortumu ile idrar ya da vajinal salgı örneğini alıp Jacobson organına götürerek koklamadan oluşur.[98] Dişi filin östrus siklüsü 14 ilâ 16 hafta arası sürer. Bunun 4 ilâ 6 haftası proliferatif dönem, 8 ilâ 10 haftası da sekretuvar dönemdir. Memelilerin çoğunda proliferatif dönemde luteinleştirici hormon bir kere artış gösterirken fillerde iki kere artış gösterir. İlk artışta dişiler erkeklere östrusta olduklarını koku yolu ile bildirir ancak ovülasyon ikinci artıştan sonra meydana gelir.[99] Dişi fillerde doğurganlık oranı 45 ilâ 50 yaşından sonra azalır.[90]
Erkek filler östrusta olan dişileri izleme ve diğer erkeklere karşı savunma davranışı gösterir. Bu davranış çoğunlukla mest hâlindeki erkek filler tarafından gösterilir ve özellikle daha yaşlı dişiler olmak üzere dişi filler bu davranışın erkek filler tarafından gösterilmesini isteyen hareketlerde bulunurlar.[100] Bu erkek fillerin üreme şansı daha yüksektir.[93] Zayıf ya da yaralı erkekler normal olarak mest olmadıkları için, mest hâli dişi fillere erkek fillerin durumunu gösterir.[101] Genç dişilere daha yaşlı erkeğin yaklaşması korkutucu gelebileceğinden aile grubundaki daha yaşlı dişiler genç dişinin yanında kalarak ona destek olur.[102] Çiftleşme sırasında erkek fil hortumunu dişinin sırtına koyar.[103] Erkek filin penisi pelvisten bağımsız olarak hereket edebilir[104] ve erkek dişinin üzerine çıkmadan önce ileri ve yukarı doğru kıvrılır. Çiftleşme 45 saniye sürer ve ne kalça hareketi ne de boşalmada duraklama görülmez.[105] Hem erkek hem de dişi filler arasında eşcinsel davranışlar oldukça sık görülür. Heteroseksüel ilişkide olduğu gibi bu davranışlarda da bir hayvan diğerinin üzerine çıkar. Erkek filler bazen oyun olarak birbirleriyle dövüşerek birbirlerini uyarırlar ve genç ile yaşlı filler arasında bağlar kurulabilir. Dişilerde eşcinsel davranışlar esaret altındaki fillerde görülür ve dişilerin birbirlerini hortumları ile tatmin ettikleri gözlemlenebilir.[106]
Doğum ve yavrular

Fillerde gebelik yaklaşık iki yıl sürer ve doğumlar arasında genelde dört ilâ beş yıllık ara bulunur. Doğumlar genelde yağmur mevsiminde yapılır.[107] Yavru filler doğduklarında yaklaşık 85 cm boyunda ve 120 kg ağırlığındadır.[102] Tipik olarak tek yavru doğar ancak ikiz doğumlara da rastlanır.[108] Görece uzun süren gebelik dönemi beş adet sarı cisim ile (çoğu memelide tektir) desteklenir ve fetüsün özellikle beyin ve hortumunun daha çok gelişmesine olanak verir.[108] Bu nedenle yaşına göre erken gelişmiş olarak doğan yavrular kısa sürede ayağa kalkıp annelerini ve aile gruplarını yürüyerek izleyebilirler.[109] Yeni doğan yavru fil aile grubundaki tüm fillerin ilgi odağıdır. Erişkinler tamamı ve genç fillerin çoğu yeni doğan yavrunun çevresine toplanıp hortumları ile ona dokunur ve okşarlar. Başlarda bir iki gün anne yavrusunun çevresinde diğer aile üyelerinin bulunmasını istemez. Bazı aile gruplarında yavrulara anneden başka dişiler bakabilir. Bu dişiler genellikle iki ilâ on iki yaş arasındadır.[102] Yakınlarda yırtıcı hayvanlar bulunduğunda aile grubu yavruları ortaya alarak toplanırlar.[110]

İlk birkaç gün yeni doğan yavru ayakları üstünde çok dengeli duramaz ve annesinin desteğine gereksinim duyar. İyi göremediği için dokunma, koklama ve işitme duyularından yardım alır. Kıvrılıp duran hortumu üzerinde çok büyük bir kontrolü yoktur ve hortumuna takılıp tökezleyebilir. İki haftalık olduğunda daha dengeli yürür ve hortumunu kontrol edebilir. Bir aylık olduğunda hortumuyla neseneleri yerden alabilir, tutabilir ve ağzına götürebilir ancak hortumuyla su çekemez. Su içmek için ağzını kullanır. Annesine hâlâ çok bağımlıdır ve yanından ayrılmaz.[109]
İlk üç ay beslenmek için tamamen annesinin sütüne bağlı olan yavru fil üç aydan sonra besin olarak bitkileri yemeye ve hortumuyla su toplamaya başlar. Aynı zamanda dudak ve bacak koordinasyonunda gelişmeler görülür. Yavru filler altı yaşına kadar süt emer ve altı aydan sonra beslenme yönünden bağımsızlık kazanırlar. Dokuz aylık olduklarında ağız, hortum ve ayak koordinasyonları mükemmelleşmiştir. Bir yaşından sonra yavru filin kendini temizleme, su içme ve beslenme yetenekleri tamamen gelişir. Yine de bir yıl kadar daha beslenme ve yırtıcılardan korunma için annesinin korunmasına ihtiyacı olur. Bir yaşından küçük yavrular saatte 2 ilâ 4 dakika kadar süt emer. Fil yavruları üç yaşına kadar süt emmeye devam eder. İki yaşından sonra süt emmeleri büyüme hızlarına, vücut kondisyonuna ve üreme yeteneklerinin gelişmesine yardımcı olur.[109] Fil yavruları arasında oyun davranışları cinsiyetlerine göre değişiklik gösterir. Dişi yavru filler birbirlerini kovalarken erkek yavru filler oyun olarak dövüş taklidi yaparlar. Dişi filler dokuz yaşında cinsel olgunluğa erişirken[102] bu süre erkek filler için 14 ilâ 15 yıl kadardır.[93] Fillerin yaşam süresi uzundur ve 60 ilâ 70 yaşına kadar yaşarlar.[60] Lin Wang adlı esaret altında yaşayan fil 86 yaşına kadar yaşamıştır.[111]
İletişim

Dokunma filler arasında önemli bir iletişim biçimidir. Bireyler birbirlerini selamlamak için hortumlarını diğerinin hortumuna dolar ve okşar. Yaşlı filler, genç filleri terbiye etmek için hortumlarıyla şaplak atar, genç filleri tekmeler ya da iter. Her yaştan ve cinsiyetten bireyler bir araya geldiklerinde ve uyarıldıklarında diğerinin ağzına, temporin bezlerine ve cinsel organlarına hortumlarıyla dokunurlar. Bu şekilde bireyler kimyasal mesajları algılayabilirler. Dokunma ayrıca anne ile yavru arasında da önemli bir iletişim yöntemidir. Hareket esnasında ana filler yavrularına hortumlarıyla, yan yana iseler ayakları ile, yavru arkadaysa kuyrukları ile dokunur. Yavru eğer dinlenmek istiyorsa annesinin ön bacaklarına dayanır. Eğer süt emmek istiyorsa annesinin göğsüne ya da bacağına dokunur.[112]
Görsel nümayiş daha çok agonistik davranışlarda ortaya çıkar. Filler kafalarını kaldırarak ve kulaklarını yanlara açarak daha tehditkâr görünmeye çalışırlar. Ayrıca nümayiş esnasında kafalarını sallayıp kulaklarını çırpar ve toz ile bitkileri hortumları ile fırlatırlar. Yapmacık kuvvet gösterisinde bulunurlar. Uyarılmış filler hortumlarını kaldırabilir. Başeğen filler kafalarını ve hortumlarını aşağı indirirken kulaklarını boyunlarına doğru yapıştırır. Meydan okumayı kabul eden filler ise kulaklarını V şeklinde açar.[113]
Filler genellikle gırtlaktan olmak üzere, bir kısmı da hortum tarafından değişikliğe uğrayan çeşitli sesler çıkarırlar. Belki de bu seslerin en bilineni, uyarıldıklarında, tehlike altında olduklarında ya da saldırı esnasında çıkardıkları boru benzeri sestir.[114] Dövüşen filler kükrer ve haykırır. Yaralı olan filler ise böğürür.[115] Hafif uyarıldıklarında uğultu gibi düşük frekanslı titreşimlerden oluşan[116] ve kısmen infrasonik sesler çıkarırlar.[117] İnfrasonik çağrılar hem Asya hem de Afrika filleri için özellikle uzun mesafe iletişimde önem taşır.[114] Asya fillerinde bu çağrılar 14-24 Hz frekansında, 85-90 dB düzeyindedir ve 10 ilâ 15 saniye sürer.[117] Afrika fillerinin çağrıları ise 15 ilâ 35 Hz aralığında ve 117 dB düzeylerindedir. Bu şekilde en çok 10 km civarında bir mesafede haberleşebilirler.[118]
_-_pone.0048907.s003.ogv.jpg)
Kenya'da Amboseli parkında çeşitli infrasonik çağrılar kaydedilmiştir. Birkaç saat ayrı kaldıktan sonra bir araya gelen aile üyelerinin çıkardığı selamlama uğultusu bunlardan biridir. Gruplarından ayrılmış bireylerin temas çağrıları yumuşak ve modüle olmamış seslerdir. Bunlara cevap olarak yüksek sesle başlayan sonra yumuşayan çağrıyla karşılık verilir. Aile üyelerine bulundukları yerden ayrılmak için grup lideri dişi tarafından yumuşak uğultu şeklinde "hadi gidelim" çağrısı yapılır. Mest dönemindeki erkekler, çok belirgin alçak frekanslı "motorsiklet" sesine benzeyen uğultular çıkarır. Bu çağrılara dişi filler koro hâlinde yine alçak frekanslı modüle seslerle karşılık verir. Çiftleştikten sonra östrusta olan dişi fil yüksek sesli bir çağrı yapar. Bir dişi çiftleştiğinde ailesi de heyecan çağrılarında bulunur.[116]
Fillerin yer yüzeyinde darbelerle oluşturulan titreşimler ve yer yüzünde akustik dalgalar hâlinde yayılan sismik yöntemle iletişim kurdukları bilinmektedir. Sismik sinyalleri algılamaya çalışan filin öne eğilerek ön ayaklarına yüklendikleri görülür ve bu davranışa "donma davranışı" denir. Filler sismik iletişime uyun olarak çeşitli adaptasyonlar geçirmiştir. Tabanlarında bulunan kıkırdaksı yumrular dişli balinalar ve sirenler gibi deniz memelilerinde bulunan akustik yağ tabakalarına benzerlik gösterir. Kulak kanalı çevresinde bulunan benzersiz bir büzgen kas kanalı sıkıştırarak akustik sinyallerin titreşimini azaltarak hayvanın daha fazla sismik sinyali duymasına olanak verir.[119] Fillerin sismik dalgaları çeşitli nedenlerle kullandığı gözlemlenir. Kaçan ya da saldırı taklidi yapan bir fil çok uzak mesafelerden duyulabiliecek sismik dalgalar yaratabilir.[120] Yırtıcılar nedeniyle kaynaklanan ikaz çağrısını sismik yolla alan filler savunma pozisyonuna girer ve aile grupları bir araya toplanır. Hareket ile üretilen simik dalgaların 32 km uzağa ulaşabildiği ancak ses yolu ile üretilen sismik dalgaların 16 km kadar yol alabildiği görülür.[121]
Zekâ ve bilişsellik
Filler öz farkındalığın ve bilişselliğin bir göstergesi sayılan ve bazı insansılar ile yunusların da başarılı olduğu ayna testinde kendilerini tanırlar.[122] Esaret altındaki bir dişi Asya fili ile yapılan araştırma sonucunda fillerin çeşitli görsel ve akustik ayırt edilebilen çiftleri öğrenebildiğini göstermiştir. Araştırma konusu olan fil bir yıl sonra aynı şekilde test edildiğinde benzer çiftlerle daha yüksek doğruluk yüzdesi elde etmiştir.[123] Filler alet kullandığı bilinen birkaç hayvan türünden biridir. Bir Asya filinin dalları uyarlayıp sineklik olarak kullandığı kaydedilmiştir.[124] Filler tarafından değişikliğe uğratılarak alet olarak kullanılan nesneler şempanzelerin kullandığı kadar karmaşık değildir. Genel olarak fillerin çok mükemmel bir hafızaları olduğuna inanılır. Bu inanışın altında bazı gerçeklerin olması muhtemeldir. Fillerin uzun sürelerde geniş boyutlarda alanları hatırlamalarını sağlayacak bilişsel haritaları olması mümkündür. Bireyler, aile üyelerinin bulunduğu yerleri akıllarında tutabilmektedirler.[51] Bilim insanları fillerin ne derecede duyguları olduğunu tartışmaktadır. Ölüm ile ilgili bazı ritüelleri olduğu görülmekte ve akraba olsalar da olmasalar da kendi türlerinin kemiklerine büyük bir ilgi göstermektedirler.[125] Ölmekte olan ya da ölü bir fil kendi grubu ve hatta kendi grubu dışındaki fillerin bile ilgi odağı olmaktadır.[126]
Korunma sorunları
Durumu
Fillerin dağılımı
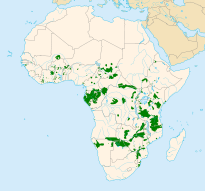
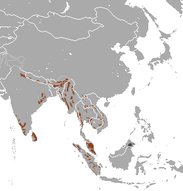
Afrika filleri, iki ırkı arasında bağımsız bir değerlendirme yapılmadan 2008 yılında Dünya Doğa ve Doğal Kaynakları Koruma Birliği (IUCN) tarafından soyunun tükenme riski yüksek olan hassas türler arasında listelenmiştir.[127] 1979 yılında Afrika'da fil popülasyonunun minimum 1,3 milyon ve maksimum 3 milyon olduğu tahmin edilmekteydi. 1989 yılında ise Afrika popülasyonu 609.000 fil olarak tahmin edilmiştir. Bunun dağılımı şöyleydi: 277.000 Orta Afrika'da, Afrika'nın doğusunda 110.000, güneyinde 204.000 ve batısında da 19.000. Daha önce tahmin edilenden çok daha az sayıda 214.000 kadar filin yağmur ormanlarında yaşadığı tahmin edilmiştir. 1987 yılından sonra fil popülasyonunun azalması hız kazanmış ve Kamerun ile Somali'de yaşayan savan ırkının sayısının %80 oranında azaldığı görülmüştür. Afrika orman ırkının sayısında ise %43 oranında azalma tespit edilmiştir. Afrika'nın güneyinde ise Zambiya, Mozambik ve Angola'da popülasyon azalırken Botsvana ve Zimbabve'de artmış, Güney Afrika'da ise sabit kalmıştır.[128] 2005 ve 2007 yılında yapılan araştırmalarda ise Afrika'nın güneyinde fil popülasyonunun yıllık %4 oranında arttığı gözlemlenmiştir.[127]
Afrika filleri en azından bulundukları her ülkede kısmen de olsa yasal koruma altındadır ancak yaşam alanlarının %70'i koruma altındaki bölgelerin dışındadır. Bazı bölgelerde başarılı koruma çabaları sonucunda yüksek popülasyon yoğunluğuna erişilmiştir. 2008 yılı itibariyle yerel popülasyonlar doğum kontrolü ve nakil yöntemleriyle kontrol altına alınmıştır. Zimbabve'nin uygulamayı bırakmasından sonra 1988'den itibaren geniş çaplı hayvan itlafı yapılmamaktadır. Afrika fili 1989 yılında Nesli Tehlike Altında Olan Yabani Hayvan ve Bitki Türlerinin Uluslararası Ticaretine İlişkin Sözleşme'nin (CITES) EK-I'inde listelenerek ticareti yasa dışı hâle getirilmiştir. 1997 yılında Botsvana, Namibya ve Zimbabve ile 2000 yılında Güney Afrika'da bulunan filler için EK-II statüsü verilmiş ve bunların ticareti sınırlı olarak belli esaslara bağlanmıştır. Bazı ülkelerde fillerin hatıra amaçlı avlanması yasaldır. Botsvana, Kamerun, Gabon, Mozambik, Namibya, Güney Afrika, Tanzanya, Zambiya ve Zimbabve gibi bu ülkeler için CITES kapsamında ihraç kotları bulunmaktadır.[127]
2008 yılında IUCN son 60-75 yıl içinde popülasyonunda %50'lik bir azalma olduğu için Asya filini soyunun tükenme riski çok yüksek olan tehlikedeki türler arasında listelemiştir.[129] CITES listesinde de EK-I altında listelenerek ticareti yasaklanmıştır.[129] Asya filleri bir zamanlar Suriye ve Irak'tan (Elephas maximus asurus alt türü) Çin'de Sarı Irmak'a ve Cava Adası'na kadar olan bölgeye dağılmışlardı.[130] Günümüzde bu bölgelerde soyu tükenmiştir[129] ve Asya filinin güncel dağılımı sınırlı bölgelerde kalmıştır.[130] Tam doğru olmasa da Asya filinin toplam popülasyonunun 40 ilâ 50 bin arasında olduğu tahmin edilmektedir. Popülasyonun yarısının Hindistan'da olması muhtemeldir. Asya fillerinin özel olarak Güneydoğu Asya'da ve genel olarak popülasyonları azalmaktaysa da Batı Gat Dağları'nda kısmen de olsa artış göstermektedir.[129]
Tehditler

Fillerin soyunun tükenmesinde en büyük tehditlerden biri dişleri, eti ve derisi için kaçak olarak avlanmasıdır.[129] Tarihsel olarak sayısız kültür, fildişinden ziynet eşyası ve diğer sanat eserleri yapmış ve hatta fildişinin kullanımı altın kullanımıyla başabaş gitmiştir.[131] Fildişi ticareti Afrika fili popülasyonunun 20. yüzyıl sonlarında azalmasında önemli bir etken olmuştur.[127] Bu nedenle, 1989 yılı Haziran ayında ABD'den başlayarak Kuzey Amerika, Avrupa ülkeleri ve Japonya gibi birçok ülkede fildişi ithalatına uluslararası yasaklamalar getirilmiştir.[131] Aynı dönemde Kenya elindeki tüm fildişi stoğunu yok etmiştir.[132] CITES, 1990 yılı Ocak ayında yürürlüğe girecek şekilde fildişi ticaretinde uluslararası yasağı onayladı.[131] Uluslararası yasakların devreye girmesinden sonra özellikle ekonomik olarak fildişi endüstrisinin büyük önem taşıdığı Hindistan ve Çin gibi ülkelerde işsizlik baş gösterdi. Aynı endüstrinin bir parçası olan Japonya ve Hong Kong ise bu duruma adapte olmayı başarabildi.[131] Zimbabve, Botsvana, Namibya, Zambiya ve Malavi fildişi ticaretine devam etmeyi istemiş ve yerel olarak fil popülasyonları sağlıklı olduğu için ve ancak doğal yollardan ölmüş ya da itlaf edilmiş hayvanların dişlerini kullanmak kaydıyla izin alabilmişlerdir.[132]
Uluslararası ticaret yasağı sayesinde Afrika'nın bazı bölgelerinde fil popülasyonu toparlanmıştır.[131] 2012 Ocak ayında Kamerun'da Bouba Njida Ulusal Parkı'nda Çad'lı baskıncılar tarafından yüzlerce fil öldürülmüştür. Bu olaylar yasakların başlamasından beri meydana gelen "en yoğun fil katli" olarak tanımlanmıştır.[132] Özellikle dişi Asya fillerinin fildişi olmadığı için, Asya filleri potansiyel olarak fildişi ticaretinden daha az etkilenir. Yine de Hindistan'da Periyar Ulusal Parkı gibi bazı bölgelerde bu tür de fildişleri için öldürülmektedir.[129]
Filler için diğer tehditler arasında yaşam alanı yok olması ve yaşam alanı parçalanması sayılabilir.[127] Asya fili, insan nüfusunun en yüksek olduğu bazı bölgelerde yaşamaktadır. Diğer simpatrik kara memelilerinden daha fazla miktarda yere gereksinim duydukları için filler insanların doğayı istila etmesinden en çok etkilenen türlerden biridir. Ekstrem durumlarda filler, insanların baskın olarak yaşadıkları bölgelerde arada kalmış küçük orman adacıklarında kısılıp kalabilmektedir. Cüsseleri ve besin gereksinimleri nedeniyle filler insanlarla birlikte tarım alanlarında bir arada yaşayamaz. Ekinleri ezen ve tüketen filler insanlarla karşı karşıya kalmaktadır. Bu nedenle yüzlerce fil ve insan geçmişte ölmüştür. Bu çatışmaların önüne geçebilmek koruma çabaları için önem arz etmektedir.[129]
Filler ve insanlar
Yük hayvanı

Filler, yaklaşık İndus Vadisi Uygarlığı zamanından[133] yani yaklaşık olarak MÖ üç binli yıllardan günümüze kadar yük hayvanı olarak kullanılmış ve kullanılmaya devam etmektedir. 2000 yılı itibariyla Asya'da 13.000 ilâ 16.500 kadar fil yük hayvanı olarak kullanılmaktaydı. Bu hayvanlar genellikle çabuk ve kolay eğitilebilecekleri bir yaş olan 10 ilâ 20 yaşlarında doğadan yakalanan hayvanlardır.[134] Eskiden geleneksel olarak tuzak ve kement ile yakalanmakta olan filler 1950'lerden beri uyuşturulmak yoluyla da yakalanmaktadır.[135] Genellikle Asya filleri yük hayvanı olarak eğitilir. Afrika'da da benzer denemelerde bulunulmuştur. Belçika Kongosu'nda Afrika fillerinin evcilleştirilmesi II. Léopold'un bir buyruğuyla 19. yüzyılda başlamıştır ve günümüzde de Api Fil Evcilleştirme Merkezi'nde devam etmektedir.[136]
Asya filleri uzak mesafelere yük taşımak, kütükleri kamyonlara doldurmak, ulusal parklarda turistleri gezdirmek, römork çekmek üzere kullanılırlar ve ayrıca dinsel geçitlerin de ayrılmaz parçasıdırlar.[134] Tayland'ın kuzeyinde Black Ivory kahvesi yapmak için kahve tanelerini sindirmeleri için kullanılırlar.[137] Görece derin sularda çalışabildikleri, çok az bakım gerektirdikleri, yakıt yerine ot ve su ile yetinebildikleri ve belirli işleri yapmak için eğitilebildikleri için mekanik aletler yerine tercih edilirler. Filler 30'dan fazla emri öğrenebilir.[134] Mest durumundaki erkek filler çalışmak için tehlikeli olabileceğinden bu durumları geçene kadar zincirlenirler.[138] Hindistan'da yük hayvanı olarak kullanılan fillerin çoğunun istismar edildiği iddia edilmiştir. Hindistan'da filler ve diğer esaret altındaki hayvanlar 1960 yılında çıkarılan Hayvanlara Zulmün Önlenmesi Yasası ile koruma altına alınmıştır.[139]
Savaş

Tarihsel olarak filler dehşet veren savaş araçları olarak görülmüşlerdir. Yanlarını korumak için zırh giydirilmiş, dişlerinin ucuna sivri demir ya da pirinç takılmıştır. Savaş filleri karşılarındaki düşman askerini yakalayıp sırtlarındaki kişiye kaldırmak ya da yere bastırıp fildişleri ile parçalamak üzere eğitilirdi.[140]
Savaş fillerine dair en eski yazılı kaynaklardan birisi MÖ 4. yüzyılda yazılmış olan ve MÖ 11. ile 8. yüzyıllar arasındaki olayları anlattığı söylenen Hint Mahabharata destanıdır. Savaş filleri Pandava ve Kaurava tarafından atlı savaş arabaları kadar sıklıkla kullanılmamıştır. MÖ 6. yüzyılda ortaya çıkan Magadha Krallığı'nda filler atlardan daha büyük kültürel önem taşımaya başlamış ve bundan sonra Hint krallıkları savaş fillerini daha yaygın olarak kullanmıştır. MÖ 5. ve 4. yüzyıllar arasında Nanda İmparatorluğu ordusunda 3.000 savaş fili varken MÖ 4. ve 2. yüzyıllar arasında Maurya İmparatorluğu ordusunda 9.000 savaş fili kullanılmış olabilir. Yaklaşık MÖ 300 yıllarında yazılmış olan Hint siyaset eseri Arthashastra'da Maurya hükümetine orduda kullanılmak üzere fillerin doğal yaşam alanı olan bazı ormanlık alanların korunması ve bu filleri öldürenlerin de idam edilmesi önerilir.[141] Fillerin savaş alanlarında kullanımı Güney Asya'dan batıya doğru Persler'e[140] ve doğuya doğru Güneydoğu Asya'ya yayılmıştır.[142] Persler savaş fillerini MÖ 6. ile 4. yüzyıllar arasında Ahameniş İmparatorluğu'nda kullanmıştır.[140] Güneydoğu Asya ülkelerinde ise ilk savaş fili kullanımı MÖ 5. yüzyılda başlar ve 20. yüzyıla kadar devam eder.[142]
Büyük İskender piyadelerini filleri yaralamaları için eğitmiş ve hem Perslerle hem de Hintlerle olan savaşlarda karşı ordularda büyük paniğe neden olmuştur. İskender'in generallerinden biri olan Ptolemaios MÖ 323 yılında Mısır'ın hükümdarı olduktan sonra ordusunda Asya fillerinden oluşan birlikler barındırmıştır. Hükümdarlığa MÖ 285 yılında başlayan oğlu ve halefi II. Ptolemaios ise savaş fillerini daha güneyden, Nubiya'dan getirtmiştir. Bundan sonra savaş filleri klasik dönem boyunca Akdeniz ve Kuzey Afrika'da kullanılmıştır. Yunan kralı Pirus MÖ 280 yılında Roma'yı işgal girişiminde savaş fillerini kullanmıştır. Romalı süvarilerin atlarını korkutmuş olsa da savaş filleri savaşın kazanılmasında belirleyici olamamış ve Pirus sonunda savaşı kaybetmiştir. Kartacalı general Hannibal Romalılar ile savaşırken MÖ 217 yılında filleri Alplerden geçirmeyi başarmış ve tamamı hayatta kalmış şekilde Po Ovası'na indirebilmiştir. Ancak filler daha sonra hastalıktan kırılmışlardır.[140]
Hayvanat bahçeleri ve sirkler

Antik dönemde filler Mısır, Çin, Yunanistan ve Roma'da menajerilerde gösteriş için tutulurdu. Romalılar filleri diğer hayvanlarla ve insanlarla gladyatör gösterilerinde karşı karşıya getirirdi. Modern Çağda filler geleneksel olarak dünya üzerindeki hemen hemen bütün hayvanat bahçelerinin ve sirklerin ayrılmaz bir parçasıdır. Sirklerde filler bazı oyunlar yapmak üzere eğitilirler. En tanınmış sirk fili muhtemelen Barnum & Bailey Sirkinin en önemli yıldız gösterilerinden biri olan Jumbo'dur (1861 - 15 Eylül 1885).[143] Filler, mest durumunda olan erkeklerin idaresinin zorluğu ve dişilerin östrus sikluslarının anlaşılamaması gibi nedenlerle esaret altında zor ürerler. Modern hayvanat bahçesi ve sirklerde bulunan fillerin çoğu Asya filidir. Asya fili 1975 yılında CITES'ın EK-I'inde listelendikten sonra 1980'lerde hayvanat bahçelerindeki Afrika fili sayısı artış göstermiştir. ABD'de esaret altında yaşayan fillerin çoğu, fil popülasyonu oldukça fazla olan Zimbabve'den gelmiştir.[144] 2000 yılında dünya üzerinde hayvanat bahçeleri ve sirklerde yaklaşık 1.200 Asya fili ve 700 Afrika fili bulunmaktadır. Esaret altındaki en büyük popülasyon 370 Asya ve 350 Afrika fili ile Kuzey Amerika'dadır. Avrupa'da yaklaşı 380 Asya ve 190 Afrika fili bulunurken Japonya'da da yaklaşık 70 Asya ve 67 Afrika fili bulunur.[144]

Fillerin hayvanat bahçelerinde tutulması bazı tartışmalara yol açmıştır. Bunu destekleyenler araştırmacıların hayvanlara ulaşımını kolaylaştırdığını ve doğal alanlarını korumak dolayısıyla da türün geleceğini sağlamak için para uzmanlık sağladığı görüşlerini savunmaktadır. Bunu eleştirenler ise hayvanların hayvanat bahçelerinde fiziksel ve zihinsel strese maruz kaldıklarını söyler.[145] Fillerin kendilerini ya da hortumlarını ileri geri sallama ve yol izleme gibi stereotipik davranışlar gösterdiği kaydedilmiştir. Bu davranış Birleşik Krallık hayvanat bahçelerinde bulunan bireylerin %54'ünde görülmüştür.[146] Avrupa hayvanat bahçelerinde bulunan fillerin doğadaki fillere göre daha kısa yaşam süresine sahip olduğu anlaşılmaktadır ancak başka araştırmalar hayvanat bahçelerindeki fillerin doğal hayatlarını sürdüren filler kadar uzun yaşadığını gösterir.[147]

Fillerin sirklerde yer alması da tartışmalara yol açmıştır. ABD'nin en önemli hayvan hakları savunucusu olan Humane Society sirkleri hayvanlara kötü davranmak ve zarar vermekle suçlamıştır.[148] 2009 yılında ABD federal mahkemesinde yaptığı tanıklık esnasında Barnum & Bailey Sirki'nin CEO'su Kenneth Feld sirk fillerinin kulaklarının arkasına, çenelerinin altına ve bacaklarına ucu metal ve ankusa adı verilen üvendirelerle vurulduğunu kabul etmiştir. Feld bu uygulamaların sirk çalışanlarını korumak için gerekli olduğunu ve elektik şoku veren bir cihaz kullanan bir fil eğiticisinin cezalandırıldığını belirtmiş ancak bu uygulamaların fillere zarar verdiğini kabul etmemiştir.[149] Bazı eğiticiler filleri fiziksel cezalandırma kullanmadan eğitmeyi denemiştir. Ralph Helfer'in aralarında filler ve aslanlar da bulunan hayvanlarını eğitirken rikkat ve ödüllendirmeye dayandığı bilinmektedir.[150]
Hastalık yayma
Memelilerin çoğu gibi fillerde insanlara hastalık geçirebilir. Bunlardan biri de tüberkülozdur. 2012 yılında Fransa'da Lyon şehrinin Tete d'Or hayvanat bahçesinde iki filde bu hastalık teşhis edilmiştir. Diğer hayvanlara ve hayvanat bahçesi ziyaretçilerine tüberkülozu geçirme riski karşısında şehir yetkililerince iki hayvan için derhal ötenazi uygulanması istenmiş ancak mahkeme kararıyla bu istek geri çevrilmiştir.[151] ABD'de Tennessee'de bir fil barınağında 54 yaşındaki bir Afrika filinin sekiz işçiye tüberküloz bulaştırdığı düşünülmektedir.[152]
Saldırılar
Filler saldırgan davranış nöbetleri sergileyebilir ve insanlara karşı yıkıcı davranışlara girebilir.[153] Afrika'da, 1970'ler ve 1980'lerde uygulanan itlaf çalışmaları sonrasında bir grup genç filin köylerde evlere zarar verdiği görülmüştür. Zamanlamasından ötürü bu saldırıların öç alma amaçlı olduğu düşünülmüştür.[94][154] Hindistan'da erkek filler düzenli olarak geceleri köylere girerek evleri yıkmakta ve insanları öldürmektedir. Jharkhand'da 2000 ile 2004 yılları arasında filler yaklaşık 300 kişi öldürmüş, Assam'da ise 2001 ile 2006 arasında fillerin 239 kişi öldürdüğü bildirilmiştir.[153] Yerel halk saldırılar sırasında fillerin sarhoş olduğunu bildirmiş ancak resmî yetkililer bu açıklamanın doğru olmadığını bildirmiştir.[155][156] Yine sarhoş olduğu düşünülen filler bir kez daha 2002 Aralık ayında bir Hint köyüne saldırarak altı kişiyi öldürmüş bunun sonucunda da yerel halk yaklaşık 200 kadar fili öldürmüştür.[157]
Kültürel tasvirler
.png)
Filler Yontma Taş Devri'nden beri sanatsal olarak tasvir edilirler. Afrika'da özellikle Sahra Çölü'nde ve güneyde çok sayıda kaya resimleri ve taş üzeri kazıma desenler bulunur.[158] Uzak Doğu'da Hindu ve Budist tapınaklarında ve ibadet yerlerinde fil motifleri görülür.[159] Filleri doğrudan görmemiş kişilerin tasvir etmesi oldukça zordur.[160] Esaret altında filler tutan Antik Romalılar Tunus ve Sicilya anatomik açıdan oldukça doğru fil tasvirlerini mozaiklere işlemişlerdir. Orta Çağın başında filleri görmemiş olan Avrupalılar genellikle bunları fantastik yaratıklar olarak resmetmiştir. Genellikle gövdeleri at ya da sığır gibi, hortumları boru gibi, dişleri domuz dişi gibi tasvir edilmiş ve hatta bazıları toynaklı resmedilmiştir. Filler, Gotik kiliseleri yapan taş ustaları tarafından sıklıkla kullanılan bir motiftir. 15. yüzyıldan itibaren, Avrupa'da hüküm süren krallara hediye olarak gittikçe artan sayıda filin gelmesi üzerine, tasvirler daha doğru olmaya başlamıştır ki bunların arasında Leonardo da Vinci'nin bir deseni de bulunur. Yine de Avrupalı sanatçılar filleri daha stilize olarak resmetmeye devam etmiştir.[161] Max Ernst 1921 yılına ait Celebes adlı sürrealist resminde fili ucundan su hortumuna benzeyen bir hortum çıkan bir silo gibi tasvir etmiştir.[162]

Fil, hem Batı hem de Doğu kültürlerinde hafızayı, bilgeliği, uzun yaşamı, başarıyı simgeler ve baba figürü olarak görülür. Birçok Afrika kabilesi için fil hayvanların kralıdır. Filler bazı dinî inanışların da öznesi olmuştur. Mbutiler ölmüş atalarının ruhlarının fillerde ikâmet ettiğine inanırlar.[159] Diğer Afrika kabilelerinde de benzer inanışlar görülür ve bazıları şeflerinin öldükten sonra fil olarak yeniden dünyaya geldiğine inanırlar. MS 10. yüzyılda Igbo-Ukwu halkı liderlerini fildişleri ile birlikte gömerlerdi.[163] Fillerin dinî önemi Afrika'da yalnızca totemiktir[164] ancak Asya'da bu önem daha dikkat çekicidir. Sumatra'da filler yıldırım ile ilişkilendirilir. Benzer şekilde Hinduizm'de tüm fillerin atası olan Airavata şimşek ve gökkuşağı ile tasvir edilir ve oraj ile bağdaştırılır.[159] En önemli Hindu tanrılarından biri olan fil başlı Ganeşa, en büyük üç tanrılar Şiva, Vişnu ve Brahma ile aynı mertebededir.[165] Ganeşa yazarlar ve tüccarlar ile bağlantılıdır ve insanlara başarı getirdiğine ve insanların arzularına ulaşmalarını sağladığına inanılır.[159] Budizm'de Buda'nın insan olarak yeniden doğan bir beyaz fil olduğu söylenir.[166] İslami gelenekte Muhammed'in doğduğu yıl Fil Yılı olarak bilinir.[167] Romalılar fillerin kendilerinin dindar olduğuna inanır ve güneş ile yıldızlara taptıklarını düşünürlerdi.[159]
Fil; zürafa, su aygırı ve gergedan ile birlikte, coğrafyasında bunlara benzer hayvanlar olmadığı için Batı'nın popüler kültüründe her yerde görülebilen egzotik amblemlerden biridir.[168] ABD'nin Cumhuriyetçi Parti'sinin sembolü olarak filin kullanımı ilk olarak Thomas Nast'ın 1874 yılında Harper's Weekly dergisinde çizdiği karikatürle başlamıştır.[169] Çocuk öykülerinde karakter olarak fillere çok sık yer verilir. Genellikle örnek davranışları olan karakterler olarak betimlenirler. Değerleri ideal insan değerleridir. Rudyard Kipling'in İşte Öyle Hikayeler kitabındaki "Çocuk Fil" öyküsü, Walt Disney'nin Dumbo'su gibi birçok öyküde tek başına kalmış olan genç fillerin yakın bağları olan topluluklarına dönüşü konu alınır. İnsani özellikler verilen diğer kurgusal fil kahramanlar arasında Jean de Brunhoff'un Babar, David McKee'nin Elmer ve Dr. Seuss'un Horton karakterleri sayılabilir.[168]
 Güney Afrika'da kaya resmi.
Güney Afrika'da kaya resmi. Ptolemaios dönemine ait altın sikke.
Ptolemaios dönemine ait altın sikke. Kamerun'dan fil maskesi.
Kamerun'dan fil maskesi.
 Seramik tablet. İran, 12. yüzyıl sonu.
Seramik tablet. İran, 12. yüzyıl sonu. İtalya'da Orta Çağdan bir fresk.
İtalya'da Orta Çağdan bir fresk.
Kaynakça
- Genel
- Shoshani, J., ed. (2000). Elephants: Majestic Creatures of the Wild. Checkmark Books. ISBN 0-87596-143-6.
- --- Shoshani, J.; Shoshani, S. L. "What is an Elephant?". s. 14-15.
- --- Shoshani, J. "Comparing the Living Elephants". s. 36-51.
- --- Shoshani, J. "Anatomy and Physiology". s. 66-80.
- --- Easa, P. S. "Musth in Asian Elephants". s. 85-86.
- --- Moss, C. "Elephant Calves: The Story of Two Sexes". s. 106-13.
- --- Payne, K. B.; Langauer, W. B. "Elephant Communication". s. 116-23.
- --- Eltringham, S. K. "Ecology and Behavior". s. 124-27.
- --- Wylie, K. C. "Elephants as War Machines". s. 146-48.
- --- McNeely, J. A. "Elephants as Beasts of Burden". s. 149-50.
- --- Smith, K. H. "The Elephant Domestication Centre of Africa". s. 152-54.
- --- McNeely, J. A. "Elephants in Folklore, Religion and Art". s. 158-65.
- --- Shoshani, S. L. "Famous Elephants". s. 168-71.
- --- Daniel, J. C. "The Asian Elephant Population Today". s. 174-77.
- --- Douglas-Hamilton, I. "The African Elephant Population Today". s. 178-83.
- --- Tuttle, C. D. "Elephants in Captivity". s. 184-93.
- --- Martin, E. B. "The Rise and Fall of the Ivory Market". s. 202-07.
- --- Shoshani, J. "Why Save Elephants?". s. 226-29.
- Sukumar, R. (2003). The Living Elephants: Evolutionary Ecology, Behaviour, and Conservation. Oxford University Press. ISBN 978-0-19-510778-4.
- Kingdon, J. (1988). East African Mammals: An Atlas of Evolution in Africa, Volume 3, Part B: Large Mammals. Academic Press. ISBN 0-12-408355-2.
- Wylie, D. (2009). Elephant. Reaktion Books. ISBN 978-1-86189-397-0.
- Özel
- ↑ Nişanyan, Sevan. "Sözlerin Soyağacı - Çağdaş Türkçenin Etimolojik Sözlüğü". 4 Eylül 2014 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20140904032018/http://www.nisanyansozluk.com:80/?k=fil. Erişim tarihi: 23 Kasım 2013.
- ↑ Liddell, Henry George; Scott, Robert (İngilizce). ἐλέφας. http://www.perseus.tufts.edu/hopper/text?doc=Perseus:text:1999.04.0057:entry=e)le/fas.
- ↑ Kalb, J. E.; Mebrate, A. (1993). Fossil Elephantoids from the Hominid-Bearing Awash Group, Middle Awash Valley, Afar Depression, Ethiopia. The American Philosophical Society. s. 52-59. ISBN 0-87169-831-5.
- ↑ Kellogg, M.; Burkett, S.; Dennis, T. R.; Stone, G.; Gray, B. A.; McGuire, P. M.; Zori, R. T.; Stanyon, R. (2007). "Chromosome painting in the manatee supports Afrotheria and Paenungulata". Evolutionary Biology 7: 6. DOI:10.1186/1471-2148-7-6.
- 1 2 Ozawa, T.; Hayashi, S.; Mikhelson, V. M. (1997). "Phylogenetic position of mammoth and Steller's sea cow within tethytheria demonstrated by mitochondrial DNA sequences". Journal of Molecular Evolution 44 (4): 406-13. DOI:10.1007/PL00006160. PMID 9089080.
- 1 2 3 4 5 Shoshani, s. 38-41.
- 1 2 3 4 Shoshani, J.; Eisenberg, J. F. (1982). "Elephas maximus". Mammalian Species 182: 1-8. DOI:10.2307/3504045. JSTOR 3504045. http://www.science.smith.edu/msi/pdf/i0076-3519-182-01-0001.pdf.
- 1 2 Cranbrook, E.; Payne, J.; Leh, C. M. U. (2008). "Origin of the elephants Elephas maximus L. of Borneo". Sarawak Museum Journal. 27 Eylül 2012 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20120927014013/http://assets.panda.org/downloads/pages_from_originofelephants_in_borneofinal2oct07_2.pdf.
- ↑ Fernando, P.; Vidya, T. N. C.; Payne, J.; Stuewe, M.; Davison, G.; Alfred, R. J.; Andau, P.; Bosi, E.; Kilbourn, A.; Melnick, D. J. (2003). "DNA analysis indicates that Asian Elephants are native to Borneo and are therefore a high priority for conservation". PLoS Biol 1 (1): e6. DOI:10.1371/journal.pbio.0000006.
- 1 2 3 4 5 Laursen, L.; Bekoff, M. (1978). "Loxodonta africana". Mammalian Species 92 (92): 1-8. DOI:10.2307/3503889. JSTOR 3503889. http://www.science.smith.edu/msi/pdf/i0076-3519-092-01-0001.pdf.
- 1 2 3 Shoshani, J. (2005). "Order Proboscidea". Wilson, D. E.; Reeder, D. M. Mammal Species of the World: A Taxonomic and Geographic Reference, Volume 1 (3. bas.). Johns Hopkins University Press. s. 90-91. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ↑ Sukumar, s. 46.
- ↑ Barnes, R. F. W.; Blom, A.; Alers M. P. T. (1995). "A review of the status of forest elephants Loxodonta africana in Central Africa". Biological Conservation 71 (2): 125-32. DOI:10.1016/0006-3207(94)00014-H.
- 1 2 3 4 Shoshani, s. 42-51.
- ↑ Grubb, P.; Groves, C. P.; Dudley J. P.; Shoshani, J. (2000). "Living African elephants belong to two species: Loxodonta africana (Blumenbach, 1797) and Loxodonta cyclotis (Matschie, 1900)". Elephant 2 (4): 1-4. http://arts.anu.edu.au/grovco/Loxodonta%20cyclotis.pdf.
- 1 2 Rohland, N.; Malaspinas, Anna-Sapfo; Pollack, Joshua L.; Slatkin, Montgomery; Matheus, Paul; Hofreiter, Michael (2007). "Proboscidean mitogenomics: chronology and mode of elephant evolution using mastodon as outgroup". PLoS Biology 5 (8): e207. DOI:10.1371/journal.pbio.0050207.
- ↑ Roca, A. L.; Georgiadis, N.; Pecon-Slattery, J.; O'Brien, S. J. (2001). "Genetic evidence for two species of elephant in Africa". Science 293 (5534): 1473-77. DOI:10.1126/science.1059936. PMID 11520983.
- ↑ Eggert, L. S.; Rasner, C. A.; Woodruff, D. S. (2002). "The evolution and phylogeography of the African elephant inferred from mitochondrial DNA sequence and nuclear microsatellite markers". Proceedings of the Royal Society B: Biological Sciences 269 (1504): 1993-2006. DOI:10.1098/rspb.2002.2070.
- 1 2 Debruyne, R. (2005). "A case study of apparent conflict between molecular phylogenies: the interrelationships of African elephants". Cladistics 21 (1): 31-50. DOI:10.1111/j.1096-0031.2004.00044.x.
- 1 2 Rohland, N.; Reich, D.; Mallick, S.; Meyer, M.; Green, R. E.; Georgiadis, N. J.; Roca, A. L.; Hofreiter, M. (2010). David Penny. ed. "Genomic DNA Sequences from Mastodon and Woolly Mammoth Reveal Deep Speciation of Forest and Savanna Elephants". PLoS Biology 8 (12): e1000564. DOI:10.1371/journal.pbio.1000564. PMC 3006346. PMID 21203580. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3006346.
- 1 2 Ishida, Y.; Oleksyk, T. K.; Georgiadis, N. J.; David, V. A.; Zhao, K.; Stephens, R. M.; Kolokotronis, S.-O.; Roca, A. L. (2011). William J. Murphy. ed. "Reconciling apparent conflicts between mitochondrial and nuclear phylogenies in African elephants". PLoS ONE 6 (6): e20642. DOI:10.1371/journal.pone.0020642.
- ↑ "Loxodonta cyclotis (Matschie, 1900)". UNEP-WCMC Species Database. http://www.unep-wcmc-apps.org/isdb/Taxonomy/tax-species-result.cfm?displaylanguage=ENG&Genus=%2525Loxodonta%2525&source=animals&speciesNo=71831&Country=&tabname=names.
- ↑ Eggert, L. S. Eggert, J. A. Woodruff, D. S. (2003). "Estimating population sizes for elusive animals: the forest elephants of Kakum National Park, Ghana". Molecular Ecology 12 (6): 1389-1402. DOI:10.1046/j.1365-294X.2003.01822.x. PMID 12755869. http://labs.biology.ucsd.edu/woodruff/pubs/210.pdf.
- ↑ Debruyne, R.; Van Holt, A.; Barriel, V.; Tassy, P.; (2003). "Status of the so-called African pygmy elephant (Loxodonta pumilio (NOACK 1906)): phylogeny of cytochrome b and mitochondrial control region sequences". Comptes Rendus de Biologie 326 (7): 687-97. DOI:10.1016/S1631-0691(03)00158-6. PMID 14556388. http://regis.cubedeglace.com/wp-content/uploads/2009/02/debruyne_et_al_20032.pdf.
- ↑ Gheerbrant, E. (2009). "Paleocene emergence of elephant relatives and the rapid radiation of African ungulates". Proceedings of the National Academy of Sciences of the United States of America (PNAS) 106 (26): 10717-10721. DOI:10.1073/pnas.0900251106. PMC 2705600. PMID 19549873. http://www.pnas.org/content/106/26/10717.full.
- ↑ Gheerbrant, E.; Sudre, J.; Cappetta, H. (1996). "A Palaeocene proboscidean from Morocco". Nature 383 (6595): 68-71. DOI:10.1038/383068a0.
- ↑ Sukumar, s. 13-16.
- 1 2 3 Shoshani, J.; Walter, R. C.; Abraha, M.; Berhe, S.; Tassy, P.; Sanders, W. J.; Marchant, G. H.; Libsekal, Y.; Ghirmai, T.; Zinner, D. (2006). "A proboscidean from the late Oligocene of Eritrea, a "missing link" between early Elephantiformes and Elephantimorpha, and biogeographic implications". Proceedings of the National Academy of Sciences 103 (46): 17296-301. DOI:10.1073/pnas.0603689103.
- 1 2 3 4 Sukumar, s. 16-19.
- ↑ Sukumar, s. 22.
- ↑ Sukumar, s. 24-27.
- ↑ Sukumar, s. 44.
- 1 2 Sukumar, s. 28-31.
- ↑ Sukumar, s. 36-37.
- 1 2 3 4 5 6 7 8 9 10 11 Shoshani, J. (1998). "Understanding proboscidean evolution: a formidable task". Trends in Ecology and Evolution 13 (12): 480-87. DOI:10.1016/S0169-5347(98)01491-8.
- ↑ Carpenter, K. (2006). Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878. Foster, J.R. ve Lucas, S.G. (eds.). "Paleontology and Geology of the Upper Jurassic Morrison Formation". New Mexico Museum of Natural History and Science Bulletin (New Mexico Museum of Natural History and Science) 36: 131-138.
- 1 2 3 4 5 West, J. B. (2002). "Why doesn't the elephant have a pleural space?". Physiology 17 (2): 47-50. PMID 11909991. http://physiologyonline.physiology.org/content/17/2/47.full.
- ↑ Debruyne, R.; Barriel, V.; Tassy, P. (2003). "Mitochondrial cytochrome b of the Lyakhov mammoth (Proboscidea, Mammalia): new data and phylogenetic analyses of Elephantidae". Molecular Phylogenetics and Evolution 26 (3): 421-34. DOI:10.1016/S1055-7903(02)00292-0. PMID 12644401. http://regis.cubedeglace.com/wp-content/uploads/2009/02/debruyne_et_al_20031.pdf.
- ↑ Noro, M.; Masuda, R.; Dubrovo, I. A.; Yoshida, M. C.; Kato, M. (1998). "Molecular phylogenetic inference of the woolly mammoth Mammuthus primigenius, based on complete sequences of mitochondrial cytochrome b and 12S ribosomal RNA genes". Journal of Molecular Evolution 46 (3): 314-26. DOI:10.1007/PL00006308. PMID 9493356.
- ↑ Gross, L. (2006). "Reading the evolutionary history of the Woolly Mammoth in its mitochondrial genome". PLoS Biology 4 (3): e74. DOI:10.1371/journal.pbio.0040074.
- ↑ Sukumar, s. 46-47.
- ↑ Choi, C. (2011). "Woolly Mammoths Could Be Cloned Someday, Scientist Says". Live Science. 1 Ekim 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20151001193228/http://www.livescience.com/17386-woolly-mammoth-clone.html.
- 1 2 3 4 Sukumar, s. 31-33.
- ↑ Vartanyan, S. L., Garutt, V. E., Sher, A. V. (1993). "Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic". Nature 362 (6418): 337-40. DOI:10.1038/362337a0.
- ↑ Tikhonov, A.; Agenbroad, L.; Vartanyan, S. (2003). "Comparative analysis of the mammoth populations on Wrangel Island and the Channel Islands". Deinsea 9: 415-20. ISSN 0923-9308.
- 1 2 Shoshani, s. 68-70.
- ↑ Somgrid, C.. "Elephant Anatomy and Biology: Skeletal system". Elephant Research and Education Center, Department of Companion Animal and Wildlife Clinics, Faculty of Veterinary Medicine, Chiang Mai University. 11 Haziran 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150611094944/http://www.asianelephantresearch.com:80/about-elephant-anatomy-and-biology-p1.php.
- ↑ Kingdon, s. 11.
- ↑ Somgrid, C.. "Elephant Anatomy and Biology: Special sense organs". Elephant Research and Education Center, Department of Companion Animal and Wildlife Clinics, Faculty of Veterinary Medicine, Chiang Mai University. 7 Haziran 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150607085533/http://www.asianelephantresearch.com:80/about-elephant-anatomy-and-biology-p4.php.
- ↑ Yokoyama, S.; Takenaka, N.; Agnew, D. W.; Shoshani, J. (2005). "Elephants and human color-blind deuteranopes have identical sets of visual pigments". Genetics 170 (1): 335-44. DOI:10.1534/genetics.104.039511. PMC 1449733. PMID 15781694. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1449733.
- 1 2 Byrne, R. W.; Bates, L.; Moss C. J. (2009). "Elephant cognition in primate perspective". Comparative Cognition & Behavior Review 4: 65-79. DOI:10.3819/ccbr.2009.40009. http://www.elephanttag.org/professional/Publications/Byrne_Bates_Moss_2009.pdf.
- 1 2 3 4 5 6 7 Shoshani, s. 78-79.
- ↑ Narasimhan, A. (2008). "Why do elephants have big ear flaps?". Indian Academy of Sciences. http://www.ias.ac.in/resonance/July2008/p638-647.pdf.
- ↑ Reuter, T.; Nummela, S.; Hemilä, S. (1998). "Elephant hearing". Journal of the Acoustical Society of America 104 (2): 1122-23. DOI:10.1121/1.423341. http://roadecology.ucdavis.edu/%5C/pdflib/Winter2005/Rodwell_Reuter_ele_ear.pdf.
- 1 2 3 4 Shoshani, s. 74-77.
- 1 2 Martin, F.; Niemitz C. (2003). ""Right-trunkers" and "left-trunkers": side preferences of trunk movements in wild Asian elephants (Elephas maximus)". Journal of Comparative Psychology 117 (4): 371-79. DOI:10.1037/0735-7036.117.4.371. PMID 14717638.
- 1 2 Sukumar, s. 149.
- 1 2 Kingdon, s. 9.
- ↑ Cole, M. (14 Kasım 1992). "Lead in lake blamed for floppy trunks". NewScientist. http://www.newscientist.com/article/mg13618470.700-lead-in-lake-blamed-for-floppy-trunks-.html.
- 1 2 Shoshani, s. 70-71.
- 1 2 3 Shoshani, s. 71-74.
- ↑ Sukumar, s. 120
- ↑ Clutton-Brock, J. (1986). A Natural History of Domesticated Mammals. British Museum (Natural History). s. 208. ISBN 0-521-34697-5.
- ↑ "Elephants Evolve Smaller Tusks Due to Poaching". Environmental News Network. 21 Kasım 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20151121185117/http://www.enn.com/wildlife/article/29620.
- ↑ Zhuoqiong, W. (16 Temmuz 2005). "Tuskless elephants evolving thanks to poachers". China Daily. http://www.chinadaily.com.cn/english/doc/2005-07/16/content_460623.htm.
- ↑ Gray, R. (20 Ocak 2008). "Why elephants are not so long in the tusk". The Daily Telegraph. http://www.telegraph.co.uk/science/science-news/3322455/Why-elephants-are-not-so-long-in-the-tusk.html.
- 1 2 Shoshani, s. 66-67.
- 1 2 3 4 Shoshani, s. 69-70.
- 1 2 3 Weissengruber, G. E.; Egger, G. F.; Hutchinson, J. R.; Groenewald, H. B.; Elsässer, L.; Famini, D.; Forstenpointner, G. (2006). "The structure of the cushions in the feet of African elephants (Loxodonta africana)". Journal of Anatomy 209 (6): 781-92. DOI:10.1111/j.1469-7580.2006.00648.x. PMC 2048995. PMID 17118065. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2048995.
- ↑ Shoshani, s. 74.
- ↑ Pennisi, E. (22 December 2011). "Elephants Have a Sixth 'Toe'". ScienceMag.org. 9 Haziran 2013 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20130609090008/http://news.sciencemag.org/sciencenow/2011/12/elephants-have-a-sixth-toe.html?ref=hp.
- ↑ Hutchinson, J. R.; Schwerda, D.; Famini, D. J.; Dale, R. H.; Fischer, M. S. Kram, R. (2006). "The locomotor kinematics of Asian and African elephants: changes with speed and size". Journal of Experimental Biology 209 (19): 3812-27. DOI:10.1242/jeb.02443. PMID 16985198.
- ↑ Genin, J. J. Willems, P. A.; Cavagna, G. A.; Lair, R.; Heglund, N. C. (2010). "Biomechanics of locomotion in Asian elephants". Journal of Experimental Biology 213 (5): 694-706. DOI:10.1242/jeb.035436. PMID 20154184.
- ↑ Hutchinson, J. R.; Famini, D.; Lair, R.; Kram, R. (2003). "Biomechanics: Are fast-moving elephants really running?". Nature 422 (6931): 493-94. DOI:10.1038/422493a. PMID 12673241.
- ↑ Shoshani, s. 60.
- ↑ Benedict, F. G.; Lee, R. C. (1936). "The heart rate of the elephant". Proceedings of the American Philosophical Society 76 (3): 335-41. JSTOR 984548.
- ↑ "How elephants 'snorkel'". BBC News. 31 Ağustos 2002. 5 Eylül 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150905143604/http://news.bbc.co.uk/2/hi/science/nature/2225846.stm.
- ↑ Shoshani, s. 80.
- 1 2 3 Eltringham, s. 124-27.
- ↑ Siegel, J.M. (2005). "Clues to the functions of mammalian sleep". Nature 437 (7063): 1264-1271. http://www.nature.com/nature/journal/v437/n7063/full/nature04285.html.
- ↑ Sukumar, s. 159.
- ↑ Hoare, B. (2009). Animal Migration: Remarkable Journeys in the Wild. University of California Press. s. 58-59. ISBN 0-520-25823-1.
- 1 2 Shoshani, s. 226-29.
- ↑ Campos-Arceiz, A.; Blake, S. (2011). "Mega-gardeners of the forest - the role of elephants in seed dispersal". Acta Oecologica 37 (6): 542-53. DOI:10.1016/j.actao.2011.01.014. http://faculty.washington.edu/timbillo/Readings%20and%20documents/CO2%20and%20Forests%20readings/Campos%20Arceiz%202011%20Elephant%20seed%20dispersal.pdf.
- ↑ Campos-Arceiz, A.; Traeholt, C.; Jaffar, R.; Santamaria, L.; Corlett, R. T. (2012). "Asian tapirs are no elephants when it comes to seed dispersal". Biotropica 44 (2): 220-27. DOI:10.1111/j.1744-7429.2011.00784.x.
- ↑ "Elephants kill endangered rhino". BBC News. 14 Şubat 2000. 18 Mayıs 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150518114712/http://news.bbc.co.uk/2/hi/642731.stm.
- ↑ Power, R. J.; Shem Compion, R. X. (2009). African Zoology 44 (1): 36-44. DOI:10.3377/004.044.0104.
- ↑ Sukumar, s. 121.
- 1 2 3 Sukumar, s. 175-79.
- 1 2 Kingdon, s. 53.
- ↑ Harris, M.; Sherwin, C.; Harris, S. (2008). "Defra final report on elephant welfare". University of Bristol. 24 Kasım 2014 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20141124052942/http://randd.defra.gov.uk/Document.aspx?Document=WC05007_7719_FRP.pdf.
- ↑ McComb, K.; Shannon, G.; Durant, S. M.; Sayialel, K.; Slotow, R.; Poole, J.; Moss, C. (2011). "Leadership in elephants: the adaptive value of age". Proceedings of the Royal Society B: Biological Sciences 278 (1722): 3270-76. DOI:10.1098/rspb.2011.0168. http://www.lifesci.sussex.ac.uk/cmvcr/Publications_files/ElephantLeadersProofs.pdf.
- 1 2 3 4 5 Sukumar, s. 179-83.
- 1 2 Highfield, R. (17 Şubat 2006). "Elephant rage: they never forgive, either". The Sydney Morning Herald. http://www.smh.com.au/articles/2006/02/16/1140064206413.html.
- 1 2 Sukumar, s. 100-08.
- ↑ Sukumar, s. 89.
- ↑ Sukumar, s. 262.
- ↑ Sukumar, s. 98-99.
- ↑ "Elephant Reproduction Project: The Estrous Cycle of Elephants". Smithsonian National Zoo. 7 Eylül 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150907192850/https://nationalzoo.si.edu/SCBI/ReproductiveScience/ElephantBreedRepro/EstrousCycle.cfm.
- ↑ Sukumar, s. 113.
- ↑ Sukumar, s. 117.
- 1 2 3 4 Moss, s. 106-13.
- ↑ Kingdon, s. 69.
- ↑ Murray E. Fowler; Susan K. Mikota (2006). Biology, Medicine, and Surgery of Elephants. John Wiley & Sons. s. 353. ISBN 978-0-8138-0676-1. http://books.google.com/?id=oCpiZA61tyQC&pg=PA353&dq=elephant+penis+mating#v=onepage&q=elephant%20penis%20mating&f=false.
- ↑ The behavior guide to African mammals: including hoofed mammals, carnivores, primates. University of California Press. 1991. ISBN 978-0-520-08085-0. http://books.google.com/books?id=g977LsZHpcsC&printsec=frontcover#v=snippet&q=elephant%20penis%20ejaculatory&f=false.
- ↑ Bagemihl, B. (1999). Biological Exuberance: Animal Homosexuality and Natural Diversity. St. Martin's Press. s. 427-30. ISBN 1-4668-0927-2.
- ↑ Sukumar, s. 259-62.
- 1 2 Lueders, I.; Niemuller, C.; Rich, P.; Gray, C.; Hermes, R.; Goeritz, F.; Hildebrandt, T. B. (2012). "Gestating for 22 months: luteal development and pregnancy maintenance in elephants". Proceedings of the Royal Society B: Biological Sciences 279 (1743): 3687-96. DOI:10.1098/rspb.2012.1038. PMC 3415912. PMID 22719030. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3415912.
- 1 2 3 Sukumar, s. 126-29.
- ↑ Kingdon, s. 64.
- ↑ "War veteran elephant dies". BBC News. 26 Şubat 2003. 21 Ekim 2014 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20141021122538/http://news.bbc.co.uk/2/hi/asia-pacific/2800737.stm.
- ↑ Payne and Langbauer, s. 116.
- ↑ Payne and Langbauer, s. 119-20.
- 1 2 Payne and Langbauer, s. 120-21.
- ↑ Kingdon, s. 63.
- 1 2 Sukumar, s. 142-45.
- 1 2 Payne, K.B.; Langbauer, W.R.; Thomas, E.M. (1986). "Infrasonic calls of the Asian elephant (Elephas maximus)". Behavioral Ecology and Sociobiology 18 (4): 297-301. DOI:10.1007/BF00300007.
- ↑ Larom, D.; Garstang, M.; Payne, K.; Raspet, R.; Lindeque, M. (1997). "The influence of surface atmospheric conditions on the range and area reached by animal vocalizations". Journal of Experimental Biology 200 (Pt 3): 421-31. PMID 9057305. http://jeb.biologists.org/cgi/reprint/200/3/421.pdf.
- ↑ O'Connell-Rodwell, E.O. (2007). "Keeping an "ear" to the ground: seismic communication in elephants". Physiology 22 (4): 287-94. DOI:10.1152/physiol.00008.2007. http://physiologyonline.physiology.org/content/22/4/287.full.pdf+html.
- ↑ O'Connell-Rodwell C. E.; Arnason, B.; Hart, L. A. (2000). "Seismic properties of Asian elephant (Elephas maximus) vocalizations and locomotion". Journal of the Acoustical Society of America 108 (6): 3066-72. DOI:10.1121/1.1323460. PMID 11144599.
- ↑ O'Connell-Rodwell, C. E.; Wood, J. D.; Rodwell, T. C.; Puria, S.; Partan, S. R.; Keefe, R.; Shriver, D.; Arnason, B. T.; Hart, L. A. (2006). "Wild elephant (Loxodonta africana) breeding herds respond to artificially transmitted seismic stimuli". Behavioural and Ecological Sociobiology 59 (6): 842-50. DOI:10.1007/s00265-005-0136-2. http://www.sscnet.ucla.edu/anthro/bec/papers/OConnellRodwell_4.16.07.pdf.
- ↑ Plotnik, J. M.; de Waal, F. B. M.; Reiss, D. (2006). "Self-recognition in an Asian elephant". Proceedings of the National Academy of Sciences 103 (45): 17053-57. DOI:10.1073/pnas.0608062103.
- ↑ Rensch, B. (1957). "The intelligence of elephants". Scientific American 196 (2): 44-49. DOI:10.1038/scientificamerican0257-44.
- ↑ Hart, B. J.; Hart, L. A.; McCory, M.; Sarath, C. R. (2001). "Cognitive behaviour in Asian elephants: use and modification of branches for fly switching". Animal Behaviour 62 (5): 839-47. DOI:10.1006/anbe.2001.1815.
- ↑ McComb, K.; Baker, L.; Moss, C. (2006). "African elephants show high levels of interest in the skulls and ivory of their own species". Biology Letters 2 (1): 26-28. DOI:10.1098/rsbl.2005.0400. PMC 1617198. PMID 17148317. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1617198.
- ↑ Douglas-Hamilton, I.; Bhallaa, S.; Wittemyera, G.; Vollratha, F. (2006). "Behavioural reactions of elephants towards a dying and deceased matriarch". Applied Animal Behaviour Science 100 (1): 87-102. DOI:10.1016/j.applanim.2006.04.014. http://www.savetheelephants.org/files/pdf/publications/2006%20Douglas-Hamilton%20et%20al%20Behavioural%20reactions%20of%20elephants%20to%20a%20dying%20matriarch.pdf.
- 1 2 3 4 5 Blanc, J. (2008). "Loxodonta africana". 2012.2. IUCN Red List of Threatened Species version 2012.2 International Union for Conservation of Nature. 13 Haziran 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150613052204/http://www.iucnredlist.org:80/details/12392/0.
- ↑ Douglas-Hamilton, s. 178-82.
- 1 2 3 4 5 6 7 Choudhury, A.; Lahiri Choudhury, D. K.; Desai, A.; Duckworth, J. W.; Easa, P. S.; Johnsingh, A. J. T.; Fernando, P.; Hedges, S.; Gunawardena, M.; Kurt, F.; Karanth, U.; Lister, A.; Menon, V.; Riddle, H.; Rübel, A.; Wikramanayake, E. (2008). "Elephas maximus". 2012.2. IUCN Red List of Threatened Species version 2012.2 International Union for Conservation of Nature. 24 Mayıs 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150524000607/http://www.iucnredlist.org:80/details/7140/0.
- 1 2 Daniel, s. 174.
- 1 2 3 4 5 Martin, s. 202-07
- 1 2 3 Christy, B. (Ekim 2012). "Ivory Worship". National Geographic. 26 Eylül 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150926142015/http://ngm.nationalgeographic.com/2012/10/ivory/christy-text.
- ↑ Sukumar, s. 57.
- 1 2 3 McNeely, s. 149-50.
- ↑ Wylie, s. 120-23.
- ↑ Smith, s. 152-54.
- ↑ Topper, R (15 Ekim 2012). "Elephant Dung Coffee: World's Most Expensive Brew Is Made With Pooped-Out Beans". The Huffington Post. 26 Kasım 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20151126070440/http://www.huffingtonpost.com/2012/10/15/elephant-dung-coffee-black-ivory_n_1968096.html.
- ↑ Easa, s. 86.
- ↑ Bist, S. S.; Cheeran, J. V.; Choudhury, S.; Barua, P.; Misra, M. K.. "The domesticated Asian elephant in India". Regional Office for Asia and the Pacific. 1 Haziran 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150601060613/http://www.fao.org:80/docrep/005/ad031e/ad031e0g.htm.
- 1 2 3 4 Wylie (2000), s. 146-48.
- ↑ Sukumar, s. 59-64.
- 1 2 Griffin, B (2004). "Elephants: From the Sacred to the Mundane". Gin Ooi, K.. Southeast Asia: A Historical Encyclopedia, from Angkor Wat to East Timor, Volume 1. s. 487-89. ISBN 1-57607-770-5.
- ↑ Shoshani, s. 168-69.
- 1 2 Tuttle, s. 184-88.
- ↑ Sterm, A. (28 Şubat 2005). "Elephant deaths at zoos reignite animal debate: Zoo supporters cite conservation, activists cite confined spaces". MSNBC/Reuters. http://www.elephants.com/media/MSNBC_2_28_05.htm.
- ↑ Harris, M.; Sherwin, C.; Harris, S. (10 Kasım2008). "Defra Final Report on Elephant Welfare". University of Bristol. 24 Kasım 2014 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20141124052942/http://randd.defra.gov.uk/Document.aspx?Document=WC05007_7719_FRP.pdf.
- ↑ Mott, M. (11 Aralık 2008). "Wild elephants live longer than their zoo counterparts". National Geographic News. 4 Mayıs 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150504050854/http://news.nationalgeographic.com:80/news/2008/12/081211-zoo-elephants.html.
- ↑ "Circus Myths: The true cruelty under the big top". Humane Society of the United States. 25 Eylül 2009. 5 Eylül 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150905065353/http://www.humanesociety.org/issues/circuses_entertainment/facts/circus_myths.html.
- ↑ Pickler, N. (4 Mart 2009). "Circus CEO says elephants are struck, but not hurt". Associated Press. http://www.dailyrecord.com/article/20090304/UPDATES01/903040429/Circus-CEO-says-elephants-struck-not-hurt.
- ↑ Wylie, s. 142.
- ↑ Anon. (2 Mart 2013). "Victory for Brigitte Bardot as elephants are reprieved". The Telegraph. http://www.telegraph.co.uk/news/worldnews/europe/france/9897992/Victory-for-Brigitte-Bardot-as-elephants-are-reprieved.html.
- ↑ Ghianni, T. (18 Şubat 2011). "Elephant behind TB outbreak at Tennessee sanctuary". Reuters. http://www.reuters.com/article/2011/02/18/us-elephant-tuberculosis-idUSTRE71H01J20110218.
- 1 2 Huggler, J. (12 Ekim 2006). "Animal Behaviour: Rogue Elephants". Londra: The Independent. http://www.independent.co.uk/environment/animal-behaviour-rogue-elephants-419678.html.
- ↑ Siebert, C. (8 Ekim 2006). "An Elephant Crackup?". Nytimes.com. 3 Kasım 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20151103001410/http://www.nytimes.com/2006/10/08/magazine/08elephant.html?pagewanted=print.
- ↑ "India elephant rampage". BBC News. 24 Aralık 1998. http://news.bbc.co.uk/1/hi/world/south_asia/241781.stm.
- ↑ "Drunken elephants trample village". BBC News. 21 Ekim 1999. http://news.bbc.co.uk/1/hi/world/south_asia/482001.stm.
- ↑ "Drunk elephants kill six people". BBC News. 17 Aralık 2002. http://news.bbc.co.uk/2/hi/south_asia/2583891.stm.
- ↑ Wylie, s. 63-65.
- 1 2 3 4 5 McNeely, s. 158-60.
- ↑ Kingdon, s. 31.
- ↑ Wylie, s. 83-84.
- ↑ Klinsöhr-Leroy, C.; Grosenick, U. (2004). Surrealism. Taschen. s. 50. ISBN 3-8228-2215-9.
- ↑ Wylie, s. 79.
- ↑ Sukumar, s. 87.
- ↑ Sukumar, s. 64.
- ↑ Sukumar, s. 62.
- ↑ Haykal, M. H. (2008). The Life of Muḥammad. Islamic Book Trust. s. 52. ISBN 978-983-9154-17-7.
- 1 2 Van Riper, A. B. (2002). Science in Popular Culture: A Reference Guide. Greenwood Press. ISBN 0-313-31822-0.
- ↑ "Cartoon of the Day: "The Third-Term Panic"". HarpWeek. 29 Mayıs 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150529193813/http://www.harpweek.com:80/09Cartoon/BrowseByDateCartoon.asp?Year=2003&Month=November&Date=7.
Dış bağlantılar
- Filler hakkında genel bilgiler içeren İngilizce bir site
- Asya filleri hakkında bilgiler içeren İngilizce bir site