Evrim
| Biyoloji alt dalı |
| Evrimsel biyoloji |
|---|
 |
|
Ana Maddeler
|
|
Tarihçe
Genel bakış
Rönesans ve Aydınlanma Darwin Öncesi Darwin • Türlerin Kökeni Sentezden Önce Modern evrimsel sentez Moleküler evrim • Evo-devo Güncel Araştırmalar |
|
Sosyal Çıkarımlar
|
|
Diğer Alanlar
|
|
Evrimsel biyoloji portalı Kategori • İlgili başlıklar • Kitap |
Evrim, biyolojide canlı türlerinin nesilden nesile kalıtsal değişime uğrayarak ilk halinden farklı özellikler kazanması sürecidir. Bazen dünyanın evrimi, evrenin evrimi gibi kavramlardan ayırmak amacıyla organik evrim ya da biyolojik evrim olarak da adlandırılır. Evrim, modern biyolojinin temel taşıdır.[1] Bu teoriye göre hayvanlar, bitkiler ve Dünya'daki diğer tüm canlıların kökeni kendilerinden önce yaşamış türlere dayanır ve ayırt edilebilir farklılıklar, başarılı nesillerde meydana gelmiş genetik değişikliklerin bir sonucudur.[1]
Evrim, bir canlı popülasyonunun genetik kompozisyonunun rastgele mutasyonlar yoluyla[2] zamanla değişmesi anlamına gelir.[3] Genlerdeki mutasyonlar, göçler veya çeşitli türler arasında yatay gen aktarımları sonucu türün bireylerinde yeni veya değişmiş özelliklerin (varyasyonların) ortaya çıkması, evrim sürecini yürüten temel etmendir. Evrim, bu yollarla oluşan değişimlerin popülasyon genelinde daha sık veya daha nadir hale gelmesiyle işler.
Dünya'daki canlı türlerinden henüz sadece 2 milyondan biraz fazlası tanımlanabilmiş ve sınıflanabilmiştir. Bazı tahminlere göre henüz tanımlanmamış 10 ila 30 milyon canlı türü vardır.[1] Bir milimetrenin binde birinden kısa bakterilerden, yerden yüksekliği 100 metreyi, ağırlığı binlerce tonu bulan sequoia servi ağaçlarına kadar dünyadaki canlı türleri, cüsse, biçim ve yaşayış biçimi açısından çok büyük farklılıklar gösterirler. Sıcak su kaynaklarında kaynama sıcaklığına yakın derecelerde yaşayan bakteriler olduğu gibi, Antarktika'daki buzullarda ya da tuz göllerinde -23 °C'ye varan sıcaklıklarda yaşayan algler ve mantarlar vardır. Aynı şekilde karanlık okyanus tabanlarındaki hidrotermal çatlakların kenarlarında yaşayan devasa boru kurtçukları olduğu gibi, Everest Dağı'nın yamaçlarında, 6 bin metre yükseklikte yaşayan hezaren çiçekleri ve örümcekler vardır.[1]
Neredeyse sınırsız sayıdaki bu çeşitli yaşam biçimleri, evrimsel sürecin bir sonucudur. Tüm canlılar, ortak atalardan geldikleri için akrabadır. İnsan ve diğer tüm memeliler, yaklaşık 150 milyon yıl önce yaşamış sivrifaremsi bir canlıdan evrimleşmiştir. Memeliler, kuşlar, sürüngenler, iki yaşamlılar ve balıkların ortak atası 600 myö yaşamış su solucanlarıdır. Tüm hayvanlar ve bitkiler, yaklaşık 3 milyar yıl önce yaşamış bakterimsi mikroorganizmalardan türemiştir.[1] Biyolojik evrim, canlı nesillerinin ortak atadan değişerek türeme sürecidir.[4] Yeni nesiller, eski nesillere göre farklılıklar taşır ve ortak atadan uzaklaştıkça çeşitlilik artar.
Evrim, biyoloji biliminin yanı sıra koruma biyolojisi, gelişim biyolojisi, ekoloji, fizyoloji, paleontoloji ve tıp gibi bilim dallarınca da başvurulan ve öğretilen bir bilimdir.[5] Bunun yanında tarım, antropoloji, felsefe ve psikoloji gibi bazı alanları da etkilemiştir. Evrimsel biyologlar evrimin bir olgu olduğunu gösteren verileri belgelerler, nedenlerini açıklayan kuramları test ederler ve geliştirirler. Bu anlamda evrim ve evrimsel süreçlerin araştırılması evrimsel biyolojinin konusudur. Evrimsel biyoloji bilimi, yaşam tarihini, onun bütünlüğüne ve çeşitliliğine yol açan evrimsel süreçler ile mekanizmaları araştırarak yukarıda sayılan disiplinlerin yanında moleküler biyoloji, davranış ve biyocoğrafya alanlarında da yapılan çalışmalara ve bu konudaki olgu ve fenomenlere ışık tutar.[6] Böylece tarihsel verilere ve adaptasyonlara dayalı açıklamalarla bu disiplinlerdeki biyolojik mekanizmalara dair yapılan çalışmaları tamamlayarak bütünleyici bir rol oynar.[6] Biyolojik bilimler genelinde evrimsel bakış açısı, gözlemler düzenleme ve yorumlama ve tahminler yapmak için genellikle vazgeçilmez ve yararlı bir çerçeve sağlar.[6] ABD Ulusal Bilimler Akademisi raporunda da (1991) vurgulandığı gibi biyolojik evrim; "modern biyolojinin en önemli anlayışı, canlıların temel yönlerini anlamak için önemli bir kavram" olarak nitelendirilir.[6]
Tarihçe
İnsanlık tarihi boyunca değişik kültürler, insanın, diğer canlıların ve evreninin kökenini çeşitli şekillerde açıklamaya çalışmış bu çaba da pek çok farklı yaratılış mitine yol açmıştır. Yahudilik, Hristiyanlık ve İslam'da canlıların ortaya çıkışı bir yaratıcının tüm evreni yoktan (Latince: ex nihilo) var etmesiyle açıklanır.
İlk Hıristiyan din adamlarından Nenizili Gregor ve Augustinus, tüm canlıların tanrı tarafından yaratılmadığını, bir kısmının sonradan tanrının yaratıklarından gelişerek oluştuğunu ileri sürmüştür. Bu iddiayı harekete geçiren güdü ise biyolojik değil, dinîdir. Bu din adamları, tüm canlı türlerinin, Tufan esnasında Nuh'un gemisine sığamayacağını, bu nedenle bir kısmının sonradan ortaya çıkmış olması gerektiğini düşünüyorlardı.[1]
Antik Yunan filozofları, kendi yaratılış mitlerini oluşturmuşlardır. Anaksimandros, hayvanların şekil değiştirebildiklerini ileri sürmüştür. Empedocles, hayvanların, önceki hayvanların organlarının birleşiminden oluştuklarını ileri sürmüştür.[1]
El-Cahiz'in Abbasiler döneminde yazdığı Hayvanlar Kitabı adlı kitapta, hayvanların evrim geçirdiği ortaya atılmıştır.
Bir olgunun ortaya çıkışında bileşenlerin değişime uğramaları ile ilgili süreç tanımının felsefi açıdan "evrim" kelimesi ile belirginleşmesi çok eskiye dayanır. Darwin'in "Türlerin Kökeni" adlı eserinde yer alan "evrimsel hayat ağacı", canlı evriminin anlatımında kullandığı mitolojik bir simgedir ve pek çok inançta yer alır. Herhangi bir "sağlam ve doğru" biyolojik altyapısı olmasa da, Aristoteles'ten Konfüçyüs'e kadar birçok önemli isim evrim kavramı konusunda yazmıştır. Ayrıca, evrim konusunda İbn-i Haldun ve İbn-i Sina farklı teoriler sunmuşlardır.
19. yüzyılda Lamarck, kazanılan karakterlerin kalıtımına dair bir hipotez öne sürmüş, fakat yaptığı deneyler bu hipotezin yanlış olduğunu göstermiştir. Aynı yüzyılda Charles Darwin, Galapagos Adaları'ndaki gözlemlerine dayanarak, evrimin mekanizmasını doğal seçilimle açıklamıştır.
Charles Darwin

Evrimin mekanizmasının anlaşılmasında ve açıklanmasında bugün geçerli olan bilimsel sentez, İngiliz doğa tarihçisi Charles Darwin tarafından 1859'da ortaya atılmış olan evrim kuramı üstüne kuruludur. Darwin, organizmaların evrim sonucu ortaya çıktığını ve organizmaların göz, kanat, böbrek gibi belirli bir amaca hizmet eden organlara sahip olmalarının yine evrimin bir sonucu olduğunu ileri sürdü. Bu iddiası temelde doğru olmakla birlikte eksikti.[1]
Darwin, kuramını doğal seçilim adını verdiği sürece dayandırıyordu. Ona göre türdeşlerine göre daha çok işe yarar özelliklere sahip olan canlılar (örneğin daha keskin görüşe sahip olanlar ya da daha hızlı koşanlar) hayatta kalma yarışında avantajlı duruma geçiyor, bu nedenle soyunu devam ettirme şansını artırıyordu.
Darwin 1831-1836 yılları arasını, işi gereği, dünyanın farklı bölgelerine seyahat ederek geçirmişti. Bu yıllarda aklında bir tür evrim kuramı şekillenmeye başladı. Farklı bölgelerde geçen 3 yıl sonunda, evrim teorisine en çok katkıda bulunacak yer olan Galapagos Adaları'na vardı. Bu adalardaki doğal yaşamı ve canlıları, Güney Amerika'dakiler (anakara) ile kıyasladı ve o dönem için şaşırtıcı bazı bağlantıları keşfetti.
Darwin burada, "başarılı nesiller sonunda, yeni bir türün, halihazırdaki bir türden yavaşça farklılaşarak oluştuğu" kanısına vardı. Doğal seçilim adını verdiği bir işlem sonucunda bu değişimlerin ortaya çıktığına inanıyordu:
Darwin'in bu teorisi 3 ana temel üzerine oturmuştur:
- Bir canlı popülasyonunda çeşitli karakteristikler mevcuttur ve bu değişken karakteristikler popülasyondaki bireyler tarafından yeni doğanlara aktarılır.
- Canlılar ölenlerin yerine geçecek sayıdan daha fazla yavrularlar.
- Ortalamada popülasyon rakamları genelde sabit kalır, hiçbir popülasyon sonsuza kadar büyüme göstermez.
Türlerin Kökeni
30 yıldan daha fazla bir süre, Darwin düşünceleri için delil topladı. 1858'e kadar fikirlerini yayımlamaktan kaçındı. Fakat 1858'de, Alfred Russel Wallace, Darwin'e Darwin'in düşüncelerine çok benzer bir evrim teorisi fikrini mektupla yollayınca, Darwin düşüncelerini kamuya sunmaya karar verdi. Daha sonra Darwin ve Wallace evrim teorisi ve doğal seçilim üzerine beraberce bir tez yazıp yayımladılar. Yine de, özellikle 1859'da yayımladığı ünlü kitabı "On The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life" (Yaşam Mücadelesinde Doğal Seçilim veya Avantajlı Irkların Muhafazası Yoluyla Türlerin Kökeni Üzerine) sayesinde Darwin'in adı Wallace'dan çok daha fazla duyuldu. Darwin'in bu kitabı daha sonra biyoloji tarihinin en etkili ve önemli kitaplarından olmuştur.
1930'lar ve sonrasında, neredeyse bir asır önce Gregor Mendel tarafından ortaya konmuş olan kalıtım kuramı, moleküler biyoloji'nin kalıtımın moleküler temellerine dair sağladığı bilgi ve Darwin'in kuramının bütünleştirilmesiyle evrim kuramı modern halini aldı. Güncel bakış açısıyla evrim, bir gen havuzu içinde bir nesilden diğerine belli bir karakterin oluşmasında etkili olan allellerden birinin sıklığının değişmesi olarak tanımlanabilir. Doğal seçilim, genetik özelliklerin üremeye katkısı, ve popülasyon yapısı bu değişime etki eden faktörlerdir. Bu güncellenmiş evrim teorisinin adı "Sentetik evrim kuramı"´dır. Sentetik evrim kuramı´nın bugünkü bilimsel değeri hakkında kuramsal biyoloji uzmanı Theodosius Dobzhansky şöyle demiştir:
| “ | "Evrimin ışığıyla aydınlatılmadıkça, biyolojide hiçbir şey bir anlam ifade etmez!" [7] | ” |
Tarihsel gelişimi
Aşağıdaki zaman çizelgesi evrimsel araştırma tarihine genel bir bakış sunmaktadır.


.jpg)





| Tarih | Bilimci | Gelişmeler |
|---|---|---|
| 1543 | Andreas Vesalius | Galen'in eserlerini incelemeye başlayan Vesalius, onun insanın anatomik yapısı hakkında verdiği bilgilerin hatalar içerdiğini görerek o zamana dek yazılmış en büyük bilim kitaplarından biri olan “De humani corporis fabrica libri septem” yani “İnsan Vücudunun Yapısı Üzerine Yedi Kitap” isimli eseriyle anatomi dalında yeni bir gelenek başlattı.[8] |
| 1628 | William Harvey | Kanın kalpten vücuda pompalandığını ve kapalı bir döngüde dolaştığını keşfeden İngiliz hekim Harvey bunu "Exercitatio Anatomica de Motu Cordis et Sanguinis in Animalibus" isimli 72 sayfalık eserinde açıklayarak yaklaşık iki yüz yıl İngiliz düşüncesini etkileyecek olan ve akıl yoluyla Tanrının planının ve tasarımının anlaşabileceğini öne süren Doğal teoloji isimli bir fikir akımının pekişmesini sağladı.[9] |
| 1666 | Nicolas Steno | Bir köpekbalığı fosilini teşrih eden Steno, köpekbalığının dişlerinin o bölgede taş parçalarıyla aynı olduğunu, daha doğrusu taş sandıklarının aslında fosil olduğunu görmüştür. Bu, paleontolojinin doğuşu olarak söylenebilir. Steno, daha sonra kayalarda araştırmalara başlamış ve aslında kayalarda yatay fosil katmanları oluştuğunu görmüştür.[10] |
| 1735 | Carl Linnaeus | Linnaeus, biyoloji ve botanikte sınıflandırma esasını getirmiş, bütün canlıları bir cetvelde göstermiştir. Onun bu metodu, bugün de kullanılmaktadır.[11] |
| 1749 | Comte de Buffon | Buffon, dünyayı yorumlayabilmesi için Yerküre tarihinin İncil’de yer almayan bir açıklaması olması gerektiğini ve dünya tarihini Isaac Newton’un yeni fiziğini kullanarak anlaması gerektiğini fark etti. Bunun yanında Buffon, "Histoire naturelle générale et particulière" isimli eserinde yaşamın belli koşullar altında kendiliğinden ortaya çıkmış olabileceğini de açıkladı. Buffon’un düşünceleri pek çok önemli gelişmeye sebep olacaktır. Cuvier’in nesil tükenmeler ile ilgili keşfi, Lyell’in dünyanın uzun bir yaşı olduğunu söylemesi ve hatta Darwin’in evrim teorisi Buffon’un görüşlerinin devamı niteliğindedir.[10][12] |
| 1789 | Thomas Malthus | Malthus, nüfus artışı hakkında yaptığı bir çalışması olan "A An Essay on the Principle of Population as it affects the Future Improvement of Society" isimli eserinde, uygun şartlarda herhangi bir popülasyonun, besin maddelerinin artışından daha hızlı bir oranda artığını ve böylece zamanla kişi başına düşen besin miktarının azaldığını öne sürerek insan davranışları için doğa bilimcilerin hayvan ve bitki popülasyonları için kullandıkları aynı prensiple hareket etmiş ve böylece Malthus'un düşünceleri daha kendisi hayattayken büyük tartışmalara neden olmuştur.[13] |
| 1812 | Georges Cuvier | Bilimsel paleontolojinin kurucusu olan Fransız doğa bilimci Cuvier, "Recherches sur les ossements fossiles de quadrupèdes" (Dört Ayaklıların Fosil Kemikleri Üzerinde Araştırmalar) isimli eserinde ve diğer çalışmalarında şaşırtıcı bir fikir öne sürerek bir zamanlar yaşamış olan bazı türlerin yerküre yüzeyinden silindiğini ve fosillerin kesik, kesik değil birbirlerinin devamı olacak şekilde olduklarını öne sürerek ilk defa "kitlesel yok oluş" fikrini ortaya koymuştur. Bu bulgular, Cuvier'e ait Doğal afet hipotezi veya Katastrofizm düşüncesini oluşturmuştur.[14] Cuvier'in Doğal Afetler Hipotezi evrimci görüşleri çağrıştırsa da evrimi açıklamada gerçek bir teori değildir.[15] |
| 1815-1822 | Jean-Baptiste Lamarck | Darwin'den önce türlerin zaman içerisinde yeni türlere değiştiğini, bugün söylediğimiz gibi evrildiğini, ilk öneren doğa bilimci Lamarck olmuştur. Lamarck'a ait olan Lamarckizm ya da diğer adıyla ”edinilmiş özelliklerin iletimi”, tüm canlıların ortak bir kökenden geldiğini ve canlının yaşadığı ortamda meydana gelen çevresel bir değişikliğin, bu ortama uymaya çalışan canlı türünün tüm (veya çoğu) üyelerinde bir değişikliğe neden olduğu ileri sürer. Örneğin Lamarck’a göre, kullanılan organlar gelişiyor, kullanılmayan organlar ise köreliyordu. Yeni kazanılan bu özellik ise gelecek nesillere kalıtım ile aktarılabiliyordu.[16] Bu durum da canlıların türleşmesine ve türlerin değişimine yol açıyordu. Lamarck’ın kalıtım düşüncesi geniş oranda popülerliğini sürdürmüş olsa da daha sonra genlerin keşfi ile bu fikir büyük oranda terk edildi.[17] |
| 1838-1859 | Charles Darwin | "Türlerin Kökeni" isimli yayınladığı ünlü kitabında Charles Darwin ve eş zamanlı olarak Alfred Russell Wallace, "Doğal Seçilim Yoluyla Evrim Teorisi"ni ortaya atmışlar ve tüm canlıların ortak bir kökenden geldiklerini, canlı türlerinin değişime uğramasının ve çeşitlenmesinin sebebinin Lamarck’ın öne sürdüğü gibi çevre değişiklikleriyle kazanılan özelliklerin ve becerilerin gelecek nesillere kalıtım yoluyla aktarılması değil, herhangi bir türün bireyleri içinde zaten var olan farklılıklar ve değişkenliklerden, bu özelliklere sahip bireylerin çevre şartlarına daha iyi uyum gösterebilenlerin diğerlerinden daha elverişli şartlar bularak daha çok üreyip çoğalabilmesiydi. Wallace ve Darwin’e göre, çevreye uyum gösterebilme ve adaptasyon, seleksiyonun bir sonucuydu, Lamarck’a göreyse bu durum çevreye uyum ihtiyacının sonucuydu.[18] |
| 1853 | Gregor Mendel | Bezelyelerin kalıtımı üzerine çalışmalar yürüten Mendel, kalıtsal özelliklerin nesilden nesile aktarıldığını kaydedip kalıtım kuramının bütün canlılar için geçerli olduğunun saptanmasına önayak oldu. Gerçi her ne kadar Mendel’in keşfinin önemi kendi zamanında fark edilmemiş olsa da daha sonra Mendel genetiği biyolojinin temel ilkelerinden biri haline gelmiştir.[19] |
| 1930 | Fisher, Haldane ve Wright | İngiliz genetikçiler ve evrimci biyologlar olan Ronald Fisher, J.B.S. Haldane ve Sewall Wright Populasyon genetiğinin kurucuları olup özellikle Fisher'in "The genetical theory of natural selection Evolution" isimli çalışmasında doğal seçilimin Mendel dünyasında nasıl işlediğini göstermede evrimin karmaşık matematiksel modellerini inşa ettiler.[20] |
| 1937 | Theodosius Dobzhansky | Ukraynalı genetikçi ve evrimsel biyolog Dobzhansky, "Genetics und the Origin of Species" (“Genetik ve Türlerin Kökeni”) isimli kitabında mutasyonların her zaman doğal olarak ortaya çıktığını, mutasyonların belli durumlarda zararlıyken, şaşırtıcı sayıda mutasyonun herhangi bir etkiye sahip olmadığını kanıtlayıp Evrimin Çağdaş Sentezini şekillendirerek, evrimsel biyoloji konusunda merkezi bir rol oynamıştır.[21] |
| 1942 | Ernst Mayr, Julian Huxley | Darwin'in Evrim Kuramı ile Mendel'in kalıtım kuramı, modern moleküler biyoloji ve matematiksel popülasyon genetiği ışığında Ernst Mayr ve Julian Huxley tarafından "Systematics and the Origin of Species" ve "Evolution - The Modern Synthesis" isimli eserlerde birleştirilerek modern evrimsel sentez adını aldı. Modern sentezin ana katkısı kalıtımın ve dolayısıyla evrimin temel birimi olan genler üzerine yeni edinilen bilgilerle evrimin mekanizması, yani doğal seçilim arasındaki bağlantıyı kurmuş olmasıdır.[22][23] |
Kalıtım
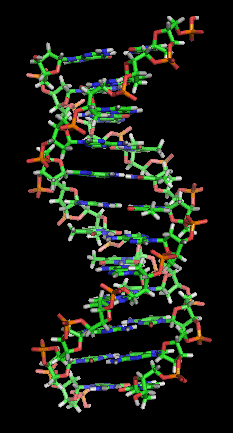
Canlılarda evrim, bir organizma için ayırt edici olan kalıtımsal fenotipik özelliklerin değişmesiyle oluşur. Örneğin, insanlarda göz rengi, kalıtsal bir özellik olup bir birey, "kahverengi göz özelliğini" ebeveynlerinin birinden miras almış olabilir.[24] Kalıtım yoluyla devralınan özellikler, genler tarafından kontrol edilir ve bir organizmanın genomu içindeki tüm gen dizilerine onun genotipi denir.[25]
Bir organizmanın yapısını ve davranışlarını oluşturan gözlemlenebilir tüm gen dizisine ise onun fenotipi denir. Bu özellikler, organizmanın sahip olduğu genotipin doğal çevre ile etkileşmesi sonucu oluşur.[26] Sonuç olarak, bir canlının sahip olduğu fenotipin birçok yönü kalıtsal olarak devredilmez. Örneğin, bronz cilt, bir kişinin genotipi ile güneş ışığı arasındaki etkileşimi sonucu meydana gelir. Böylece, insanlarda bronz cilt, çocuklara kalıtım yoluyla aktarılmaz. Ancak, bazı insanlar, genotiplerindeki farklılıklar nedeniyle diğerlerine göre daha kolay bronzlaşır. Çarpıcı bir örnek, bronzlaşmanın görülmediği ve güneş yanığına karşı çok hassas olunan albinizm özelliğinin kalıtsal olarak aktarılabilmesidir.[27]
Kalıtsal özelliklerin, genetik bilgiyi kodlayan bir molekül olan DNA yoluyla nesilden nesile aktarıldığı bilinmektedir.[25] DNA, dört çeşit bazdan oluşan uzun bir polimerdir. Belli bir DNA molekülü boyunca sıralanmış baz dizileri, bir cümleyi belirleyen harf dizileri gibi, genetik bilgileri belirler. Bir hücre bölünmeden önce DNA kopyalanır ve böylece ortaya çıkan her iki hücre, DNA dizisini kalıtım yoluyla devralır. Tek bir işlevsel birimi belirleyen DNA molekül parçalarına gen denir. Farklı genler, farklı baz dizilerine de sahiptir. Uzun DNA iplikçikleri, hücre içinde kromozom adı verilen, yoğunlaşmış yapılar oluşturur. Bir DNA dizisinin kromozom içindeki belirli konumuna ise lokus denir. Eğer bir lokus üzerindeki DNA dizisi, bireyler arasında farklılık gösteriyorsa, bu dizinin farklı formlarına da alel denilir. DNA dizileri, yeni aleller üreten mutasyonlar tarafından değiştirilebilirler. Bir gen içinde mutasyon oluştuğunda, ortaya çıkan yeni alel, canlının fenotipini değiştirerek geni düzenleyen veya kontrol eden özelliğe etki edebilir.[3] Ancak, bir alel ile bir özellik arasındaki bu basit iletişim, bazı durumlarda işlerken, çoğu özellikler daha karmaşık olup çoklu etkileşen genler tarafından kontrol edilirler.[28][29]
Son bulgular, DNA içindeki nükleotid dizileri değişimleri ile açıklanamayan kalıtsal değişikliklere dair önemli örneklerin olduğunu doğrulamıştır. Bu fenomenler, epigenetik kalıtım sistemleri olarak sınıflandırılmıştır.[30] Kromatinleri imleyen DNA metilasyonu, kendi kendini idame ettiren metabolik döngüler, RNA enterferansı karışımı ile genlerin susturulması ve proteinlerin üç boyutlu uyarlaması (örneğin prion gibi), epigenetik kalıtım sistemlerinin organizma düzeyinde keşfedildiği alanlardır.[31][32] Gelişimsel biyologlar, genetik ağlardaki karmaşık etkileşimlerin ve hücreler arasındaki iletişimin, gelişimsel plastisite ve genetik kanalizasyondaki bazı mekanizmaların desteklediği kalıtsal varyasyonlara yol açabileceğini göstermektedir.[33] Kalıtsallık, daha büyük ölçeklerde de oluşabilir. Örneğin, niş oluşturma süreci ile edinilen ekolojik miras, organizmaların doğal çevrelerindeki düzenli veya belirli aralıklarla tekrarladıkları etkinlikler olarak tanımlanır. Bu, gelecek nesillerin ekosistemdeki hiyerarşide konumlarını seçmek veya değiştirmek için bir miras oluşturur. Gelecek nesil, böylece, onların genlerini ve atalarının daha önce gerçekleştirdiği ekolojik faaliyetler ile oluşan çevresel özellikleri devralır.[34] Evrimde, doğrudan genlerin kontrolü altında olmayan diğer kalıtım olaylarına örnekler, simbiyoz oluşumları açıklayan simbiyogenez ile kültürel özelliklerin kalıtıldığı ve "gen-kültür ortak evrimi" olarak da bilinen ikili kalıtımdır.[35][36]
Genetik varyasyonların oluşumu


Bir canlı organizmanın fenotip özellikleri, onun sahip olduğu genotip ile içinde yaşadığı çevrenin etkisine dayanır. Bir popülasyon içindeki fenotiplerdeki değişimin önemli bir kısmı, genotipler arasındaki farklılıklardan kaynaklanır.[37] Modern evrimsel sentez, evrimi, bu genetik varyasyonlarda zaman içinde oluşan değişimler olarak tanımlar. Bir genin belirli bir alel frekansı, aynı genin diğer form ve tiplerine oranla, zaman içinde daha çok veya daha az yaygın hale gelirler. Yeni bir alel tipi, sabitleşme noktasına geldiğinde, ya ondan önce gelen alel tipinin yerini alarak ya da tamamıyla popülasyondan silinerek, bir varyasyonun kaybolmasına yol açar.
Doğal seçilim, bir popülasyonda sadece yeteri kadar genetik varyasyonlar var olduğunda evrime neden olur. Mendel genetiğinin keşfinden önce, karışmalı kalıtım isimli hipotez, yaygın bir görüştü. Karışmalı kalıtım teorisine göre, doğan yavrular, ebeveynlerinin özelliklerinin ortalamasını taşıyacaktı. Buna göre, eğer birisinin anne ve babasından biri uzun boylu, öbürü kısa boylu ise, kendisi orta boylu olacaktır. Ancak, karışmalı kalıtım doğru olmuş olsaydı, genetik varyasyonların hızla kaybolmaları gerekiyordu ve bu da, doğal seçilim yoluyla evrimi mantıksız kılıyordu. Hardy-Weinberg Kuralı ise, popülasyondaki varyasyonların nasıl korunduğuna dair çözüm sunabilmiştir. Hardy-Weinberg kuralına göre, yeterince büyük bir popülasyon içinde var olan alellerin frekansları (genlerdeki varyasyonlar), yumurta ve spermin oluşumu sırasında gen parçaları alellerin rastgele karıştırılması ve üreme hücrelerinin döllenmesi sırasında bu alellerin rastgele kombinasyonları gibi popülasyona etki eden güçler tarafından sabit kılınır.[38]
Bu anlamda varyasyonlar, genetik materyallerdeki mutasyon, eşeyli üreme yoluyla genlerin yeniden karıştırılması ve popülasyonlar arasındaki göçler (gen akışı) sonucu oluşur. Mutasyonlar ve gen akışı ile popülasyonlarda sürekli yeni varyasyonlar oluşsa da, bir türün genomunun büyük bölümü, o türün tüm bireylerinde benzer olup özdeştir.[39] Buna rağmen, genotipteki nispeten küçük farklılıklar bile, fenotiplerde dramatik farklılıklara yol açabilirler. Örneğin, şempanze ve insan genomu arasındaki farkların büyük bir kısmı, % 5,07 oranında delesyon ve insersiyonlardan kaynaklanırken nükleotit farklılıklar ise sadece % 1.52 oranında olup böylece toplam sapma % 6,58 olarak tahmin edilmiştir. Hatta, DNA tekrarları ile düşük karmaşıklıktaki DNA'lar çıkarıldığında bu farklılık oranı % 2,37'ye kadar düşmektedir.[40]
Mutasyon
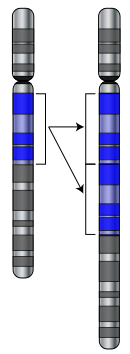
Mutasyonlar, bir hücre genomunun DNA dizisinde oluşan değişimlerdir. Mutasyonlar, her zaman organizma üzerinde negatif veya pozitif etkilere sahip olmayabilirler. Oluşan mutasyonların bir gen ürünün değişmesinde veya genin doğru ya da tamamen işlemesini engellemede herhangi bir etkileri de olmayabilir. Drosophila melanogaster sineği üzerinde yapılan çalışmalar, gen tarafından oluşturulan bir proteinin mutasyonunda, bu mutasyonun yaklaşık %70'inin zararlı etkilere sahip olduğunu, geri kalanının ise ya nötr ya da zayıf faydalı etki gösterdiğini ortaya koymaktadır.[41]
Mutasyonlar, genellikle genetik rekombinasyonlarla çoğaltılan (bkz. Gen duplikasyonu) kromozomların büyük bölümlerini içerebilirler ve böylece, bu genetik rekombinasyonlar sayesinde, genomda bir genin ek bir kopyası ortaya çıkabilir.[42] Genlerin ek olarak kopyalanması, yeni genlerin geliştirilebilmesi için gerekli olan hammaddenin önemli bir kaynağıdır.[43] En yeni genler, ortak ataları paylaşan ve kendinden önce var olmuş olan genlerin gen ailelerinden oluştuğu için bu genlerin ek olarak kopyalanması önemlidir.[44] Örneğin, insan gözü ışığı algılayan yapılar oluşturmak için dört adet değişik gen kullanır: Bu genlerden üçü, renkli görmek için ve bir tanesi ise gece görüşü için kullanılır. Bu dört genin hepsi de, tek bir atasal genden türemiştir.[45]
Atasal bir gen, bu genin bir kopyası mutasyona uğrayıp yeni bir fonksiyona sahip olduğunda, yeni tür genler oluşturabilir. Bu yöntem, sistemin yedeklenmesini artırdığı için bir genin kopyalanması durumunda daha kolaydır. Bu şekilde, gen çiftindeki bir gen, yeni bir işlev kazanırken diğer gen kopyası, eski işlevini sürdürmeye devam eder.[46][47] Diğer tip mutasyonlar ise, dana önceki şifrelenmemiş genlerden tamamen yeni genler bile oluşturabilirler.[48][49]
Yeni bir gen nesli, aynı zamanda kopyalanan birçok genlerin küçük parçalarını da içerebilir ve bu parçalarla yeni fonksiyonlara sahip yeni kombinasyonlar oluşturabilirler.[50][51] Yeni genler, daha önceki parçaların karışmasıyla bir araya getirilirken bu parçalardan oluşan kümeler, daha karışık işlevlere sahip yeni kombinasyonların oluşması için birbirleriyle karıştırılabileceği, bağımsız ve basit işlevlere sahip modüller gibi davranırlar.[52] Örneğin, poliketit sentazları, antibiyotik oluşturan büyük enzimlerdir ve montaj hattındaki her bir adım gibi, tüm süreçlerdeki her kataliz de böyle bir adım olup yaklaşık yüz kadar küme içermektedir.[53]
Eşeyli üreme ve rekombinasyonlar
Eşeysiz üreyen organizmalarda, genler, üreme sırasında başka bir bireyin genleriyle karışmadığı için birlikte bağlanırlar ve kalıtımla topluca aktarılırlar. Buna karşılık, eşeyli üreyen canlıların yavruları, ebeveynlerinin rastgele karıştırılmış ve bağımsız örneklerinden oluşan kromozomlarını içerirler. Eşeyli üreyen canlılar, homolog rekombinasyon adı verilen benzer bir süreçte, birbirleriyle uyuşan iki kromozom değiştirirler.[54] Rekombinasyonlar ve bağımsız örneklerin birbiri arasında değiş tokuş edilmesi, alel frekanslarını değiştirmezler, ancak bunun yerine, hangi alelin hangi alel ile bağlantılı olacağını değiştirerek yeni alel kombinasyonları sayesinde yeni yavrular üretirler.[55] Eşeyli üreme, genellikle genetik varyasyonları ve evrim hızını artırır.[56][57]
Gen akışı
Gen akışı, popülasyonlar ve türler arasında oluşan gen alış verişleridir.[58] Bu nedenle, gen akışı, bir popülasyon veya tür için yeni bir varyasyon kaynağı teşkil edebilir. Farelerin, kara içlerindeki popülasyonlardan kıyılardaki popülasyonlara göç etmeleri veya çiçek polenlerinin, rüzgar aracılığıyla ağır metallere karşı hassas olan bir bitki popülasyonundan ağır metallere karşı daha toleranslı olan başka bir bitki popülasyonuna (veya tersine) yol almaları gibi, bireylerin, birbirinden ayrı bulunan farklı canlı popülasyonlar arasındaki bu gibi gidiş gelişleri, göç ve hareketleri gen akışına yol açabilir.
Türler arasındaki bu gen transferi, hibrit canlıların oluşumunu ve yatay gen transferini de içerir. Yatay gen transferi, bir canlının, onun yavrusu olmayan başka bir canlıya, genetik materyal transfer etmesidir. Bu, bakteriler arasında en yaygın olanıdır.[59] Tıpta bu durum, antibiyotik direnci kazanan bakterilerin, bunu diğer türlere hızla aktardığı ve bu şekilde bakteriler arasında bu rezistans genlerin yayılması olayında görülür.[60] Genlerin yatay transferi, bakterilerden ökaryotlara doğru olmuştur. Bir maya türü olan Saccharomyces cerevisiae ile fasulye böceği Callosobruchus chinensis arasında olduğu gibi, bakterilerden ökaryotlara, iki değişik alem arasında da yatay gen transferleri olmuştur.[61][62] Daha büyük ölçekte gerçekleşen yatay gen transferlerine örnek, genlerindeki dizileri bakteriler, mantarlar ve bitkiler gibi üç ayrı alemden alan ökaryotik Bdelloidea türü rotiferalardır (tekerlekli hayvan).[63] Aynı zamanda virüsler de gen transferi ile farklı biyolojik alemler içindeki canlılar arasında DNA taşıyabilirler.[64]
Büyük ölçekli gen transferleri, ökaryotik hücreler ile bakterilerin ataları arasında gerçekleşen kloroplast ve mitokondri devralımı sırasında da gerçekleşmiştir. Ökaryotların kendilerinin, bakteriler ile arkealar arasında gerçekleşen yatay gen transferi sonucu oluşmuş olmaları da mümkündür.[65]
Evrimi oluşturan süreçler
Evrimi sürdüren iki temel süreç vardır: Doğal seçilim ve genetik sürüklenme. Bu süreçlerin ilki olan doğal seçilim, bulunduğu ortama en iyi uyum sağlayan bireylerin hayatta kalmasını ve kendi genlerini döllerine aktarmasını, rakip bireylerin ise üreme şansı bulamayıp genlerinin ortadan kalkması sonucunu doğurur. Doğal seçilim ile hayatta kalmaya yardımcı olan yeni özellikler sağlayan mutasyonlara sahip bireyler hayatta kalarak popülasyonda baskın hale gelir, hayatta kalma şansını azaltan mutasyonlara sahip bireyle ise yok olur. Bu sayede sonraki nesildeki bireyler, atalarından aldıkları genler sayesinde ortama daha iyi uyum sağlar ve hayatta kalmakta daha başarılı olurlar.[3][66] Çok sayıda nesil sonrasında, çok sayıda başarılı, küçük, rastgele değişikliğin birikmesi ile adaptasyonlar belirgin hale gelir, bu sayede türler çevrelerine olası en iyi uyumu sağlamış olurlar.[67]
İkinci temel süreç ise genetik sürüklenmedir. Genetik sürüklenme, popülasyonda genlerin görülme sıklığında rastgele değişimlere yol açar. Bir nesilde görülen rastgele bir genetik sürüklenme, daha sonraki nesillerde birikim sağlayarak organizmada belirgin değişimlere yol açar.
Evrim, aynı türden canlıların birbirleriyle karıştığı bir popülasyonda, alel frekanslarının değişimleri sonucu meydana geldiğinden [38] doğal seçilim ve genetik sürüklenmenin yanında, genetik otostop , mutasyon ve gen akışı gibi, alel frekanslarının değişimine yol açan diğer mekanizmaların olduğu da bilinmektedir.
Doğal seçilim
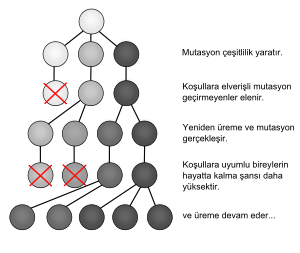
Evrime göre canlılığın devamı ve çeşitliliği doğal seçilimle sağlanır. Doğal seçilimin üç temel bileşeni bulunur: Genetik karakterlerin devamını sağlayan kalıtım, farklı karakterlerin popülasyondaki zenginliğini sağlayan çeşitlilik, ve bu çeşitli karakterlerden doğadaki koşullara en uygun olanının hayatta kalmasını sağlayan seçilim.
Bu temellere göre Darwin, her popülasyonda birçok bireyin hayatta kalamadığı, kurtulamadığı veya üreyemediğini belirtmiştir. Varolma mücadelesinde sınırlı birçok kaynak için ve mevcut riskler (yırtıcı hayvanlar vb.) yüzünden popülasyonun her bireyi bir diğeriyle yarışmaktadır. Bu varolma mücadelesinde, ortama en iyi adapte olabilmiş bireyler seçici bir avantaja sahip olmakta, daha çok yaşamakta ve daha çok üreyebilmektedir.
Doğal seçilim yoluyla evrim, üremeyi düzelten ve geliştiren genetik mutasyonların bir popülasyonun birbirlerini izleyen nesilleri içinde daha sık ve yaygın olarak görülmeye başlaması sürecine verilen isimdir. Çoğu zaman "apaçık" mekanizma olarak tanımlanan doğal seçilim yoluyla evrim, aynı zamanda zorunlu olarak aşağıdaki üç basit gerçeği takip eder:
- Kalıtsal varyasyonlar, halihazırda popülasyonun canlı bireyleri arasında mevcut bulunmaktadır.
- Canlılar hayatta kalabileceğinden çok daha fazla yavrular üretir.
- Bu yavruların hayatta kalma ve üreme yetenekleri de birbirlerinden farklıdır.
Bu koşullar, canlılar arasında hayatta kalma ve üremeye dair rekabete yol açar. Sonuç olarak, rakiplerine karşı kendilerine avantaj sağlayan özelliklere sahip bireyler bu özellikleri gelecek nesillere aktarırken bir avantaj kazandırmayan özelliklerse gelecek nesillere aktarılmazlar.[68]
Doğal seçilimin ana fikri bir canlının evrimsel uyumluluğudur veya diğer bir tanımlamayla seçilim değeridir.[69] Bir canlının uyumluluğu veya seçilim değeri ise, onun gelecek nesillere olan genetik katkısının boyutunu belirleyen hayatta kalma ve üreme yeteneği ile ölçülür.[68] Ancak, seçilim değeri üreme sonrası oluşan yavruların toplam sayısı değildir: bunun yerine bir canlının genlerini taşıyan sonraki nesillerin oranı ile gösterilir.[70] Örneğin, bir canlı iyi bir şekilde hayatta kalıp sayıca hızlı üreyebildiği halde onun oluşturduğu yavrular hayatta kalabilmek için çok zayıf ve küçük ise, bu durumda bu canlı gelecek nesiller için ancak çok küçük bir değerde genetik katkı yapmış olur ve düşük seçilim değerine sahip olur.[71]
Eğer bir alel, bu genin diğer alellerinden daha çok seçilim değerini artırıyorsa o zaman bu alel her nesille birlikte popülasyon içinde daha yaygın hale gelecektir. Bu özelliklere "lehine seçilmiş" özellikler denir. Seçilim değerini artıran özelliklere örnek, daha iyi hayatta kalma ve artış gösteren doğurganlıktır. Bunun tersine, daha az yararlı ya da zararlı aleller seçilim değeri düşürür ve bu da bu alellerin daha nadir şekilde görülmesine neden olur. Buna da "aleyhine seçilmiş" özellikler denir.[72] Daha da önemlisi, bir alellin seçilim değeri sabit bir özellik değildir; çevre şartlarının değişmesi durumunda daha önce nötr veya zararlı olan özellikler faydalı, daha önce yararlı olan özellikler ise zararlı hale gelebilirler.[73] Bunun yanında, seçilim yönü bu şekilde her ne kadar ters olsa bile, geçmişte kaybedilen özellikler benzer bir şekilde tekrar evrimleşmezler (bkz: Dollo yasası).[74][75]
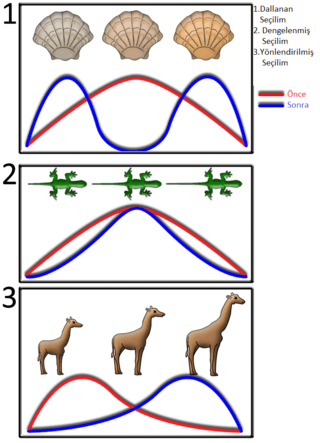
Popülasyon içinde, çeşitli değerlere sahip bir özelliğin (örnek: boy uzunluğu) doğal seçilimi üç farklı şekilde olabilir. Bunlardan biri, zaman içinde bir özelliğin ortalama değerinin değiştiği yönlendirilmiş seçilimdir. Örneğin canlıların yavaş yavaş büyüyüp öncekilerinden daha uzun boylu olmaları gibi.[76] İkincisi ise bu özelliklerin aşırı uç değerlerinin seçildiği dallanan seçilimdir ve genellikle iki farklı değerin ortalama değerden daha yaygın hale gelmesiyle sonuçlanır (iki tip veya bimodal dağılım). Bu tür bir seçilim tipinde sadece uzun boyluluk ve kısa boyluluk avantaj sağladığından canlılar da uzun boylu ve kısa boylu olarak çeşitlenmeye başlarlar. Buna karşı ortalama boylu olmak avantajını yitirdiğinden popülasyon içinde orta boya sahip canlılar azalmaya başlar. Son olarak dengelenmiş seçilim ise her iki uçtaki aşırı uç değerlerin ayıklandığı bir seçilim türüdür. Ortalama değer dışında seyreden varyanslar ve çeşitlilik bu durumda azalma göstermeye başlar.[77][78] Bu durum, örneğin, organizmaların yavaş yavaş aynı boya veya uzunluğa sahip olmasına yol açar.
Doğal seçilimin özel bir örneği, bir canlının diğer potansiyel eşler üzerisindeki cinsel çekiciliğini artırarak çiftleşme başarısını yükselten özelliklerin seçildiği cinsel seçilimdir.[79] Kullanışsız büyük boynuzlar, gürültülü çiftleşme ve kur yapma çağrıları, dikkat çekici parlak renkler her ne kadar onları avlayan hayvanların da ilgisini çekip erkek bireylerin hayatta kalma şansını azaltsa da cinsel seçilim yoluyla evrimleşmiş buna benzer özellikler, bazı hayvan türlerin erkeklerinde özellikle belirgindir.[80] Hayatta kalmaya dair dezavantaj yaratan bu tür özellikler, erkek bireylerin sahip olduğu daha yüksek üreme başarısı tarafından dengelenirler ve bu durum da cinsel seçilim yoluyla evrimleşen özelliklerin taklit edilmelerinin zor olduğunu göstermektedir (Handikap ilkesi).[81]
Doğal seçilim, hangi bireylerin ve bireysel özelliklerin daha çok veya daha az hayatta kalacağı hakkında büyük oranda genellikle doğayı ölçü alır. Bu anlamda "doğa", canlıların yaşadıkları çevrede, fiziksel ve biyolojik her türlü element ve unsurlarla etkileşim gösterdiği ekolojik sistemi kasteder. Ekolojinin kurucusu Eugene Odum ekolojik sistemi, "Belirli bir alanda fiziksel çevre ile etkileşime giren... bütün organizmaları içeren herhangi bir birim... böylece sistem içinde açıkça belirlenmiş olan trofik yapıya, biyolojik çeşitliliğe ve madde döngülerine (örneğin, canlı ve cansız birimler arasındaki madde alış verişine) yönlendirir" diyerek tanımlar.[82] Ekolojik sistem içindeki her bir popülasyon, ayrı bir niş veya sistemin diğer bölümleriyle farklı ilişkileri olduğu bir konum işgal ederler. Bu ilişkiler, canlıların yaşam öyküsünü, besin zinciri içindeki konumunu ve coğrafi olarak erişim alanını belirler. Bu geniş doğa anlayışı, doğal seçilimi kapsamı içine alan tüm belirli kuvvetlerin tanımlanmasında bilim insanlarına yardımcı olur.
Doğal seçilim, genler, hücreler, canlı birey, organizma grupları veya türler gibi bir yapının farklı seviyeleri içinde etki gösterebilir veya hareket edebilirler.[83][84][85] Bunun yanında, doğal seçilim aynı anda birden çok düzeyde de hareket edebilir.[86] Birey seviyesi altında gerçekleşen doğal seçilime bir örnek, transpozon adı verilen ve genom içinde çoğalıp yayılabilen genlerdir.[87] Aşağıda da anlatılacağı gibi, birey üzerindeki seviyede gerçekleşen seçilimler (örneğin, grup seçilimi), karşılıklı yardımlaşmanın evrilemesine izin verirler.[88]
Yapay seçilim

Yapay seçilim, insanların bilinçli olarak bir organizmanın belli özelliklerini seçmesi sürecidir. Yapay seçilim, evcil hayvan ve bitkilerin kontrollü olarak yetiştirilmesi sonucu gerçekleşir. İnsan eliyle hangi hayvan ya da bitkinin üretileceğine karar verildiğinde, hangi genlerin gelecek nesillere aktarılacağına da karar verilmiş olunur. Bu anlamda, hangi organizmanın üreyeceğine, hangi istenilir özelliklerin korunacağına doğa yerine insanlar karar verir.
Yapay seçilimin en büyük etkisi evcil hayvanlarda gözlenir. Örneğin Danua ve Çivava köpek cinslerinin arasındaki cüsse farkı yapay seçilimin bir sonucudur. Çok farklı görünmelerine rağmen, her iki köpek cinsi de -diğer tüm evcil köpek cinsleri gibi- günümüzden yaklaşık 15.000 yıl önce Çin'e denk gelen bölgede evcilleştirilmiş olan birkaç kurdun soyundan gelir.[89] Bunun gibi, bitki yetiştiriciliğinde insanlar, bir türün (örneğin buğday bitkisinin) yalnızca kendilerine daha fazla besini daha kolay şekilde sağlayan bireylerini ellerinde tutup yetiştirerek o türde evrimsel değişime yol açabilirler. Ziraatte iyi bilinen geleneksel ıslah yöntemlerinin birçoğu yapay seçilime örnektir. Yapay seçilim doğal seçilime benzer, ancak çok önemli bir fark, doğal seçilimde insanlar yerine doğanın kendisi seçme işini üstlenmiştir.[90]
Gen aktarımlı bitkiler veya genetiği değiştirilmiş organizmalar ise, modern genetik mühendislik yöntemleri ve rekombinant DNA teknolojisi ile, olumsuz çevre koşullarına karşı daha dayanıklı olması ve en az maliyetle en verimli ürünü vermeleri için bu organizmaların bazı gen bölgelerinin laboratuvar koşullarında yapay olarak değiştirildiği bitkilerdir.
Genetik sürüklenme
Genetik sürüklenme ya da "Sewall Wright etkisi", küçük bir grup canlının genetik havuzunda tamamen şans eseri oluşmuş değişikliklerdir.[91] Genetik sürüklenme bir popülasyondaki genetik bir karakteristiğin yok olmasına ya da güçlü olanın hayatta kalmasından ve alellerin değerinden "bağımsız olarak" yaygın hale gelmesine neden olur.[91] Popülasyonda üremeyi gerçekleştiren canlıların sayısı arttıkça, genetik sürüklenmenin etkisi azalır. Bu durum yazı-tura örneğine benzer. Art arda iki kere tura gelmesi doğal karşılanırken 20 kere tura gelmesi tuhaftır. Yazı-tura işlemi tekrarlandıkça, turaların oranı %50'ye yaklaşır.[1]
Genetik sürüklenmenin etkisi en çok, bir canlı türünün kaderi birkaç bireye bağlı olduğunda ortaya çıkar. Bu duruma kurucu prensibi denir. Göl, ada gibi izole olmuş ortamlara rüzgar veya başka canlıların vücudu gibi herhangi bir vasıtayla ulaşan tohumlar ve hayvan türleri, genellikle ulaştıkları yeni ortamda koloniler oluşturur. Bu birkaç kurucu bireydeki alellerin görülme sıklığı, genellikle geride bıraktıkları popülasyondaki lokusların çoğundan farklıdır. Bu farklılıklar, yeni ortamda türeyen popülasyon üzerinde uzun süreli evrimsel etkiler yaratır. Hawaii Adaları gibi takımadalarda görülen tür çeşitliliğinin, birbirine temas eden anakaralardan fazla olmasının nedeni, kurucu prensibidir.[1]
Yanlı mutasyon
Önemli bir varyasyon kaynağı olmanın yanı sıra, farklı mutasyonların oluşması için moleküler düzeyde farklı olasılıklar var olduğunda, yanlı mutasyon olarak bilinen bir süreçte, mutasyon da bir evrim mekanizması olarak işlev görebilir.[92] Eğer iki genotip, örneğin nükleotit G ile aynı pozisyondaki başka bir nükleotit A, aynı seçilim değerine sahipse ama G'den A'ya olan mutasyon, A'dan G'ye olan mutasyonlardan daha sık görülüyorsa, bu durumda A'ya sahip genotip gelişme eğiliminde olacaktır.[93] Farklı taksonlarda yanlı mutasyonların katılımı veya silinmesi farklı genom boyutlarının evrimine yol açabilir.[94][95] Gelişimsel veya mutasyonla ilgili bu tür bir yanlılık ve tarafgirlik morfolojik evrimde de gözlemlenmiştir.[96][97] Örneğin, Baldwin etkisi olarak da bilinen evrimin ' önce fenotip ' teorisine göre, mutasyonlar sonunda daha önce çevre şartları tarafından uyarılan özelliklerin genetik asimilasyonuna neden olabilirler.[98][99]
Yanlı mutasyon efekt ve etkileri diğer süreçlerin üstünü kapamıştır. Eğer her iki mutasyona da sahip olmak hiçbir ek avantaj içermemiş ve bu yüzden seçilim iki mutasyondan biri lehine olmuşsa, bu durumda popülasyon içinde daha çok sabitleşen mutasyon, aynı zamanda en sık olarak görülen mutasyon olacaktır.[100][101] Bir genin fonksiyon kaybına yol açan mutasyonlar, tam işlevlere sahip yeni bir gen oluşturan mutasyonlardan çok daha yaygındır. Fonksiyon kaybına yol açan mutasyonların çoğu buna rağmen seçilmişlerdir. Ancak, seçim zayıf olduğunda yanlı mutasyonlar fonksiyon kaybına rağmen evrimi etkileyebilirler. Örneğin, pigmentler mağarada karanlıkta yaşayan canlılar için artık yararlı olmadıklarından kaybolma eğilimi gösterirler.[102] Mutasyon yanlılığı nedeniyle veya fonksiyonlar bir bedele sahip olduğu için bu tür bir fonksiyon kaybı meydana gelebildiği gibi bir kez fonksiyon avantajı yitirildiğinde doğal seçilim kayıplara yol açabilir. Laboratuvardaki evrim sırasında bir bakteride (Bacillus subtilis) spor oluşturma yeteneğinin kaybolması, spor oluşturma yeteneğinin bedeline karşı doğal seçilimden ziyade mutasyon yanlılığı tarafından yol açılmışa benziyor. İşlev kaybına dair herhangi bir seçilim olmadığında kaybın oluştuğu hızın, etkin popülasyon büyüklüğünden ziyade,[103] mutasyon oranına bağlı olması da bu durumun genetik sürüklenmeden çok mutasyon yanlılığı tarafından desteklendiğini göstermektedir.
Genetik otostop
Remkombinasyonlar, aynı DNA dizisi üzerinde bulunan alellerin birbirlerinden ayrılmasını sağlarlar. Ancak, rekombinasyonların oranı düşüktür (her kromozom ve her popülasyon başına yaklaşık iki defa vuku bulurlar). Sonuç olarak, kromozom üzerinde birbirine yakın olan genler her zaman karıştırılarak birbirinden ayrılmazlar ve birbirlerine yakın genler, genetik bağlantı olarak bilinen bir fenomenle toplu şekilde kalıtılarak miras bırakılırlar.[104] Bu eğilim, bağlantı dengesizliği adı verilen olasılık beklentilerine göre, iki alellin hangi sıklıkta tek bir kromozom üzerinde görülmelerinin saptanmasıyla ölçülür. Genellikle bir grup içinde kalıtılan ve miras bırakılan bir alel dizisine haplotip denir. Bu, belirli bir haplotip içindeki bir alellin çok avantajlı olduğu durumlarda önemli olabilir. Doğal seçilim, haplotipteki diğer alellerin de popülasyon içinde daha yaygın hale gelmesine yol açan seçici süpürmeyi tetikleyebilir. Bu etkiye, genetik otostop ya da genetik taslak denir.[105] Genetik taslağın oluşmasına yol açan neden, bazı nötral genlerin, seçilim etkisi altında uygun bir etkin popülasyon büyüklüğü tarafından bağlanmış olan diğer genlerle genetik olarak bağlantılı olması gerçeğinden kaynaklanır.[106]
Gen akışı
Gen akışı, popülasyon ve türler arasında genlerin alış verişi veya değiş tokuş edilmesidir.[58] Gen akışının varlığı ya da yokluğu, evrimin gidişini temelden değiştirir. Organizmaların karmaşıklığı nedeniyle, zaman içinde tamamen birbirinden izole olmuş herhangi iki popülasyon, bu her iki popülasyon çevreye uyumluluk açısından temel olarak aynı kalsalar bile, sonunda Bateson-Dobzhansky-Muller Modelinde olduğu gibi nötral süreçler yoluyla genetik uyuşmazlıklar ve bağdaşmazlıklar geliştirecektir.
Eğer popülasyonlar arasında genetik farklılaşmalar baş gösterirse, popülasyonlar arasındaki gen akışı, yerel popülasyondaki dezavantajlı olan özellik ve alellerin ortaya çıkmasına yol açabileceği gibi, bu popülasyon içindeki canlıların, genetik olarak uzak popülasyonlarla çiftleşmesini önleyen ve sonunda yeni türler ortaya çıkması ile sonuçlanan mekanizmalar geliştirmesine neden olabilir.Böylece, canlı bireyler arasında genetik bilgilerin alınıp verilmesi, temel olarak biyolojik tür kavramının geliştirilmesinde önemlidir.
Modern evrimsel sentezin gelişimi sırasında Sewall Wright tarafından geliştirilen Değişken denge kuramı'nın kısmen izole olmuş popülasyonlar arasındaki gen akışı, adoptiv evrimin önemli bir parçasıydı.[107] Ancak, son zamanlarda Değişken denge kuramının önemine dair ciddi eleştiriler olmuştur.[108]
Birlikte evrim
Birlikte evrim, iki veya daha fazla canlı türünün, birbirlerinin evrimini karşılıklı olarak etkilemesidir.[109] Burada bir türün evrimi, diğer türde adaptasyonların oluşmasına yol açar. İkinci türdeki bu değişimler, daha sonra da birinci türü etkileyerek yeni adaptasyonların oluşmasına yol açar. Buna birlikte adaptasyon denir. İşte bu karşılıklı yanıt ve seçilim döngüsü, birlikte evrim olarak adlandırılır.[110] Örneğin bir bitkinin morfolojisindeki evrimsel bir değişiklik, o bitkiyle beslenen bir otçulun morfolojisini etkileyebilir. Otçulda meydana gelen değişiklik de tekrar bitkiyi etkileyebilir ve bu süreç karşılıklı devam eder.
Birlikte evrim, farklı türlerin ekolojik etkileşimleri arttığında gerçekleşme eğilimindedir. Bu ekolojik etkileşimler şöyle sıralanabilir:[109]
- Avcı - av
- Parazit - ev sahibi (konak)
- Mücadele halindeki türler
- Ortak yaşamlı türler

Birlikte evrimin en bariz örnekleri çoğunlukla ortak yaşamlı olan bitki-böcek çiftlerinde görülür. Birçok bitki ve onların polen taşıyıcıları olan böcekler varlıklarını devam ettirebilmek için birbirlerine bağımlıdırlar. Ancak polen taşıyıcısı olmayan hayvanlarla eşleşmiş bitki türleri de mevcuttur.[109]
Bazı Orta Amerika akasyaları, içi boş dikenlere ve yapraklarının sapında nektar salgılayan gözeneklere sahiptir. Acacia sphaerocephala (boğa boynuzlu akasya), dikenlerinin içine yuva yapan ve nektarla beslenen Pseudomyrmex karıncalarına ev sahipliği yapar. Karıncalar da akasyayı çeşitli otçullara karşı korur.[109] Bu ilişki birlikte evrimin bir sonucudur.[111] Bitki karıncaların barınabilmesi için içi boş dikenleri ve nektar salgılayan gözenekleri oluşturmuş, karıncalar da bitkiyi otçullardan koruyan davranış biçimini geliştirmişlerdir.[109] Karıncalar bitkiye zarar veren her türlü böcek ve tırtılı öldürmenin yanı sıra bitkinin civarındaki araziyi yabani otlardan temizlemekte, gölge yapan yakındaki ağaçlara zarar vermektedirler.[111] Boğa boynuzlu akasya ve karınca arasındaki bu ilişki ilk kez 1874'te doğa tarihçisi Thomas Belt tarafından gözlenmiştir.[111]
Diğer bir örnek, Kuzey Amerika'da yaşayan ve zehirli olarak bilinen sarı semenderin ürettiği tetrodotoksin zehiri ve onu avlayan Thamnophis sirtalis sirtalis isimli adi bahçe yılanın tetrodoksine karşı geliştirdiği dirençtir. Bu av-avcı çiftinde, evrimsel silahlanma yarışı, semenderde çok yüksek düzeyde bir zehir seviyesi ve yılanda da buna bağlı olarak çok yüksek düzeyde bir antitoksin direncine yol açmıştır.[112]
Evrimin canlılar üzerindeki etki ve sonuçları
Evrim, organizmaların şekil ve davranışlarını her yönden etkiler. En göze çarpan, kendine özgü davranışsal ve fiziksel uyarlamalar, doğal seçilimin sonuçlarıdır. Bu adaptasyonlar, besin arayışı, avcı hayvanlardan korunma, eşler üzerinde cinsel çekicilik gibi diğer etkenlerin yardımıyla seçilim değerini artırırlar. Bunun yanında, canlı organizmalar, genellikle akrabalarına yardım etme veya karşılıklı yarar getiren simbiyoz ilişkilere girme suretiyle karşılıklı yardımlaşma yaparak da seçilime tepki verebilirler. Uzun vadede, evrim, organizmaların ata popülasyonlarını, artık melez bireyler üretemeyecekleri yeni gruplara bölerek yeni türler meydana getirir.
Evrimin bu sonuçları bazen; soy tükenmesi, türleşme, bir tür veya popülasyon içindeki adaptasyonlar gibi küçük evrimsel değişmeler olan mikro evrim ile türler seviyesinde veya türler üstünde oluşan makro evrim olmak üzere ikiye ayrılır.[107] Genel olarak, makro evrim, mikro evrimin uzun dönemlerdeki sonuçları olarak kabul edilir.[113] Böylece, mikro ve makro evrim arasındaki ayrım, temel bir ayrım olmayıp bu fark sadece zamanla ilgilidir.[114] Bunun yanında, makro evrimde, tüm türlerin özellikleri önemli olabilir. Örneğin, bireyler arasındaki varyasyonların büyük bir miktarı, bir türün yeni yaşam alanlarına hızla uyum sağlamasına izin verir. Buna karşın, popülasyonun bir parçasının izole kalmasının daha olasılı hale geldiği geniş bir coğrafi alana yayılmak türleşme şansını artırırken şans ve avantajların azalması ise soy tükenmesine yol açar. Bu anlamda, mikro evrim ve makro evrim farklı seviyelerde seçilim içerirler: Bunlar, genler ve canlılar üzerinde etkin olan mikro evrim ile türlerin seçilimi gibi evrimsel süreçlerin tüm türler ile türleşme ve soy tükenme oranları üzerinde etkili olduğu makro evrimdir.[115][116]
Yaygın bir yanlış anlaşılma, evrimin amaçları ve uzun vadeli planları olduğudur. Gerçekçi olmak gerekirse, evrimin hiçbir uzun vadeli amaçları olmadığı gibi hedefi, daha büyük bir karmaşıklık üretmek değildir.[117][118] Evrimle daha karmaşık yapıya sahip canlılar meydana gelmiş olsa da bu, organizmalarının toplam sayısının artmasının bir yan etkisi olarak ortaya çıkmıştır ve hâlâ olsun biyosferde yaygınlık gösteren basit yaşam biçimleri günümüzde de değişmeden kalabilmişlerdir.[119] Örneğin, türlerin ezici bir çoğunluğu, çok küçük boyutlarına rağmen dünya biyokütlesinin yarısından çoğunu oluşturan mikroskobik prokaryotlar olup [120] yeryüzündeki biyo çeşitliliğin büyük çoğunluğunu oluştururlar.[121] Basit organizmalar, bu nedenle, tarihler boyunca yeryüzündeki hayatın baskın yaşam formu olmuşlardır ve daha hissedilebilir ve belirgin oldukları için yanıltıcı bir şekilde sadece çeşitlenmiş gibi gözüken kompleks yaşamlı günümüzde de ana yaşam formu olmaya devam etmektedirler.[122] Gerçekten de, hızlı üreme şekilleri, adaptasyonların ve evrimin eş zamanlı olarak gözlemlenmesine ve deneysel evrim üzerinde çalışmalara olanak verdiği için mikroorganizmaların evrimi, modern evrimsel araştırmalarda özellikle önem taşımaktadır.[123][124]
Adaptasyon
Adaptasyon, canlıları yaşadıkları ortama daha uygun hale getiren bir süreçtir.[125][126] Ayrıca, adaptasyon terimi, bir canlının hayatta kalması için önemli olan bir özelliği de tanımlar. Örneğin, at dişlerinin ot ve çimleri öğütecek şekildeki adaptasyonu. Adaptasyon teriminin evrimsel süreçler ile adaptif özelliklerin anlatımında kullanılması, bu sözcüğün iki ayrı anlama geldiği şekilde ayırt edilebilir. Adaptif uyarımlar, doğal seçilimin ürünüdür.[127] Aşağıdaki tanımlar Theodosius Dobzhansky'a aittir:
- Adaptasyon, bir organizmanın içinde yaşadığı ortam veya habitatlarda daha iyi yaşamasını mümkün kıldığı evrimsel süreçtir.[128]
- Uyumluluk, bir canlının, belirli bir dizi yaşam ortamlarında yaşayabilme ve üreyebilme derecesini gösteren adapte olma halidir.[129]
- Adaptif bir özellik, bir canlının hayatta kalma ve üreme olasılığını sağlayan veya artıran gelişim şeklinin bir görünümüdür.[130]
Adaptasyon, ya yeni bir özelliğin kazanılmasına ya da eski bir atasal özelliğin kaybına yol açar. Her iki türdeki değişimleri gösteren bir örnek, antibiyotik seçilimlerde ilaçların hedeflediği etkileri değiştiren veya ilacı hücre dışına pompalayan taşıyıcıların faaliyetlerini artıran genetik değişikliklerle antibiyotik direncin oluştuğu ve bakterilerin gösterdiği adaptasyonlardır.[131] Diğer çarpıcı örnekler ise, uzun vadeli bir laboratuvar deneyinde Escherichia coli bakterisinin sitrik asiti besin olarak kullanma özelliği geliştirmesidir. Flavo bakterisi, ona naylon sanayisinin yan ürünlerinden beslenip büyümesini sağlayabilen yeni ve benzersiz bir enzim geliştirmiş[132][133] ve toprakta yaşayan bir mikro organizma olan Sphingobium bakterisi ise sentetik bir pestisit olan Pentaklorofenol'u indirgeyebilen, tamamen yeni bir metabolik yöntem geliştirmiştir.[134][135] İlginç ama tartışmalı bir fikir, bazı uyarlamaların, organizmaların genetik çeşitlilik oluşturma yeteneğini artırdığını ve doğal seçilim tarafından canlıların evrilebilirliğini artırarak uyarlandığıdır.[136][137][138][139]
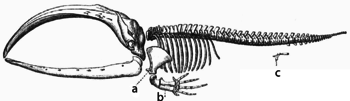
Adaptasyon, hali hazırda mevcut olan yapıların kademeli değişimleri yoluyla oluşup meydana gelirler. Sonuç olarak, benzer iç organizasyon yapıları, ilgili canlı organizmalarda farklı işlevlere sahip olabilirler. Bu, tek bir atasal yapının, farklı şekildeki işlevlere adapte olmasının bir sonucudur. Yarasa kanatları içindeki kemikler, tüm bu yapıların memelilerin sahip olduğu ortak bir atadan türemiş olmalarıyla nedeniyle, örneğin farelerin ayaklarındaki ve primatların ellerindeki kemiklere, çok benzerlik gösterirler.[141] Ancak, tüm canlı organizmalar bir ölçüde birbirleriyle akraba oldukları için,[142] eklem bacaklıların, kalamarlar ve omurgalıların gözleri ya da eklem bacaklılar ile omurgalıların bacakları ve kanatları gibi, az ya da ortak hiçbir yapısal benzerliklere sahip değilmiş gibi görünen organlar bile, derinlemesine homoloji olarak tanımlanan, işlevlerini ve bağlanımlarını kontrol eden ortak bir homolog gen üzerinde birbirleriyle bağlantılı olabilirler.[143][144]
Evrim sırasında, bazı yapılar özgün işlevlerini kaybedip körelmiş yapılara dönerler.[145] Bu tür yapılar, ata türde veya diğer yakın akraba türlerde net bir işleve sahipken mevcut türlerde az veya hiçbir işleve sahip olmayabilirler. Bunlara örnek, Psödogenler,[146], kör mağara balıklarındaki işlevini yitirmiş göz kalıntıları,[147] uçma özelliğini yitiren kuşların kanatları,[148] ve balina ile yılanlarda mevcut olan kalça kemikleridir.[140] İnsanlarda körelmiş yapılara dair örnekler ise, yirmi yaş dişleri,[149] koksiks (kuyruk kemiği),[145] apandis (vermiform appendix) [145] veya tüylerin ürpermesi[150] ile ilkel refleksler gibi diğer davranışsal izlerdir.[151][152][153][154]
Ancak, basit uyarlamalar gibi görünen pek çok özellikler, aslında yapıların başlangıçta özgün bir işlev için uyarlandığı ama süreç içinde başka bir işlev için yararlı hale geldiği ön uyarlamalardır.[155] Bir örnek, kovuk yarıklarında saklanabilmek için son derece düz ve yassı bir kafa yapısı geliştirmiş olan Holaspis guentheri Afrika kertenkelesi olup onun yakın akrabalarına bakıldığında da bu özelliğe rastlanabilinmektedir. Ancak, bu canlıda yassı kafa formu, onun ağaçtan ağaca bir planör gibi süzüp kaymasına yardımcı olacak biçimde gelişmiş olan bir ön adaptasyondur.[155] Hücreler içinde, bakteri kamçısı gibi moleküler makineler ve protein sıralama mekanizmasından sorumlu düzenekler, daha önce mevcut olan ve farklı işlevlerle yükümlü birkaç proteinin yerleştirilmesiyle evrilip gelişmişlerdir.[107] Başka bir örnek, organizmaların gözleri içindeki mercekte kristalin olarak adlandırılan ve yapısal protein olarak işlev görmeye yarayan enzimlerin, glikoliz ve ksenobiyotik metabolizmadan alınıp buraya yerleştirilmeleridir.[156][157]
Ekolojinin önemli bir prensibi, iki farklı türün ekolojik nişi sürekli olarak işgal edemeyip bir tanesinin elenmesi ilkesine dayanan Gause kuralı veya rekabetçi dışlanım ilkesidir.[158] Sonuç olarak, doğal seçilim, türleri farklı ekolojik nişlere uyum sağlama zorunluluğunda bırakan bir eğilim gösterecektir. Bu durum, örneğin, iki değişik çiklit balığı (Cichlidae) türünün, beslenme yönünden aralarındaki rekabeti en aza indireceği için farklı habitatlara uyum göstererek birbirinden farklı yaşam alanlarında yaşaması anlamına gelebilir.[159]
Evrimsel gelişim biyolojisinin güncel bir araştırma alanı, adaptasyonlar ve ön uyarlamaların gelişimsel biyolojik temelleridir.[160] Bu araştırmaların konusu, embriyonik gelişimin kökeni ve evrimi ile gelişimsel süreçlerin ve gelişimdeki değişikliklerinin nasıl yeni özellikler ürettiği ile ilgili araştırmalardır.[161] Bu çalışmalar, evrimin, memelilerdeki orta kulak kısmını oluşturmak yerine diğer hayvanlarda çene haline dönüşen embriyonik kemik yapıları gibi yeni yapıların gelişimini değiştirebileceğini göstermiştir.[162] Bunun yanında, evrim süreçlerinde gelişimden sorumlu genlerin değişmesi yüzünden, tavuklarda embriyoların timsah benzeri dişlerle büyümesine neden olan bir mutasyon gibi, kaybolan yapıların yeniden belirmesi ve ortaya çıkması da mümkündür.[163] Organizmaların yapı ve şekillerindeki birçok değişimlerin, korunmuş genlerin küçük bir diziliminde olan değişiklikler olduğu şimdi açık hale gelmektedir.[164]
Karşılıklı yardımlaşma
Türler arasında, birlikte evrilen her etkileşim şekli zorunlu olarak çekişme doğurmaz.[165] Çoğu durumda, her iki tarafa yarar getiren etkileşimler de ortaya çıkar. Örneğin, bitki kökleri üzerinde büyüyen ve topraktaki besinleri soğurup emmede bitkilere yardımcı olan mikorhizal mantarlar ile bitkiler arasında çok sıkı bir iş birliği ve karşılıklı yardımlaşma mevcuttur.[166] Bu karşılıklı ilişkide bitkiler de, fotosentez ile mantarın ihtiyaç duyduğu şekeri temin ederler. Burada, mantarlar, bitkinin bağışıklık sistemini baskılayan sinyaller göndermek suretiyle konukçu ev sahibi ile besin alış verişi yapar ve bu şekilde aslında bitki hücrelerinin içinde gelişip büyürler.[167]
Bunun gibi, aynı türün canlıları arasında da koalisyon oluşumları gelişip evrilmiştir. Kısır ve üreyemeyen böceklerin, kolonide üreme özelliği taşıyan küçük sayıdaki canlıları beslediği ve koruduğu, arılar, termitler ve karıncalar gibi sosyal yaşayan böcek türlerinde gerçek sosyal yaşam biçimi evrilmiştir. Daha küçük ölçeklerde, istikrarlı bir organizma yapısı sürdürebilmek için bir canlı hayvanın vücudunu üremede sınırlayan somatik hücreler, hayvanın küçük sayıdaki germ hücrelerini yavru oluşturabilmesinde desteklerler. Burada, somatik hücreler, onlara büyümeleri veya olduğu gibi kalmaları ya da ölmeleri için talimat verdikleri belirli sinyallere yanıt verirler. Eğer hücreler bu sinyallere aldırmazlar ve uygunsuz şekilde çoğalırlarsa, bu durumda onların kontrolsüz büyümesi kansere yol açar.[168]
Türler arasındaki bu tür bir karşılıklı yardımlaşma, bir canlının, yavrusunu yetiştirip büyütmesinde akrabasına yardımcı olduğu akraba seçilimindeki süreçler tarafından evrilmiştir.[169] Bu tür bir etkinliğin seçilmiş olmasının olası bir nedeni; eğer akrabalarına yardım eden canlı birey, yardım etme etkinliğini destekleyen ve teşvik eden genlere sahipse, onun akrabaları da muhtemelen bu genlere sahip olacak ve böylece bu aleller aktarılabilecektir.[170] Canlılar arasındaki evrimsel iş birliğini ve karşılıklı yardımlaşmayı teşvik eden diğer süreçler, bu yardımlaşmanın bir grup organizmaya yarar sağladığı grup seçilimini içerir.[171]
Türleşme
Türleşme, bir türün birbirlerinden ayrılarak gelecek nesillerde iki veya daha çok türlere ayrılması sürecidir.[172]
Tür kavramını tanımlarken birden çok yol vardır. Hangi kavramın kullanılacağı, ilgili türlerin gösterdiği özelliklerle ilişkilidir.[173] Örneğin, tür kavramlarından bazıları, eşeyli üreyen organizmaları daha kolay tanımlama özelliğine sahipken diğerleri eşeysiz üreyen canlıları tanımlamakta daha uygundur. Çeşitli tür tanımlarının olmasına rağmen, bu kavramlar, üç büyük felsefi yaklaşımdan birisine dahil edilebilirler: melezleşme, ekolojik ve filogenetik.[174] Biyolojik tür tanımı (BTT), melezleşme yaklaşımının klasik bir örneğidir. 1942 yılında Ernst Mayr tarafından tanımlanan biyolojik tür tanımına göre, tür, aralarında gen alışverişi yapan ya da bu potansiyelde olan doğal populasyon gruplarının oluşturduğu birliktir. Böyle bir populasyon, diğer populasyonlardan üreme bakımından izole olmuştur ve onlarla gen alışverişi yapamaz.[175] Geniş anlamdaki ve uzun vadedeki kullanımına rağmen, biyolojik tür tanımı da diğer tanımlar gibi, özellikle Prokaryotlarda [176] tartışmasız değildir ve bu probleme "türler sorunu" denmektedir.[177] Bazı araştırmacılar, tüm tür tanımlarını kapsayıcı ve birleştirici, monistik bir tür tanımı oluşturmayı denemişlerken, diğerleri çoğulcu bir yaklaşımı benimseyerek tür tanımının mantıksal olarak yorumlanmasında farklı yollar ve metotlar olabileceğini öne sürmüşlerdir.[173][178]
Birbirinden ayrılarak farklılaşan iki popülasyonun, birbirleriyle üremelerinin önündeki engeller (üreme yalıtımı), popülasyonların yeni türler oluşturması adına zaruridir. Gen akışı, yeni genetik varyantları diğer popülasyonlara da yayarak bu süreci yavaşlatabilir. İki türün, en son ortak atadan bu yana birbirinden ne kadar uzaklaşmış olmalarına bağlı olarak, at ve eşeklerin çiftleşerek kısır ve üreyemeyen melez katırları meydana getirmeleri gibi,[179] bu türler, yine de yeni döller ve yavrular oluşturabilirler. Bu hibritler genellikle kısırdır. Bu durumda, yakın akraba olan türler, birbirleriyle sürekli olarak çiftleşip melezleşebilirler, ancak seçilim bu melezler lehine olmayıp türler de birbirinden ayır ve farklı kalacaktır. Ancak, kimi zaman bu çiftleşmelerden yaşama yeteneğine sahip melezler de oluşur ve oluşan bu yeni türler, ya ebeveyn türler arasındaki ara özellikler gösterirler ya da tamamen yeni bir fenotipe sahip olurlar.[180] Yeni hayvan türlerinin oluşmasında melezleşmenin önemi, özellikle iyi bilinen bir örnek olarak gri ağaç kurbağasında (Hyla versicolor) olduğu gibi, birçok hayvan türlerinde görülmüş olmasına rağmen bu mekanizmaların tam olarak nasıl işlediği belirsiz olup henüz açıklığa kavuşmamıştır.[181]
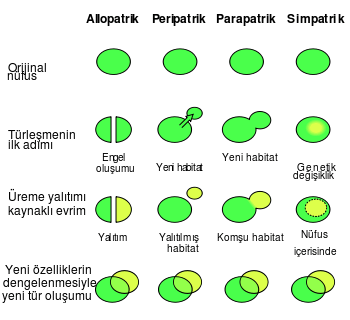
Türleşme, hem kontrollü laboratuvar koşullarında hem de doğada, birden çok defa gözlemlenmiştir.[182] Eşeyli üreyen organizmalarda türleşme, jenealojik sapmayı takiben üreme yalıtımı sonucu meydana gelmektedir. Dört farklı türleşme mekanizması vardır. Hayvanlarda en yaygın olan türleşme şekli, ilk başta, habitat parçalanması veya göç gibi nedenlerle birbirinden coğrafi olarak yalıtılan popülasyonlarda görülen allopatrik türleşme şeklidir. Bu tür koşullar altındaki bir seçilim, organizmaların görünüm ve davranışlarında çok hızlı değişiklikler meydana getirebilirler.[183][184] Seçilim ve sürüklenme, kendi türlerinin geri kalanından izole edilmiş popülasyonlar üzerine bağımsız olarak etki ederlerken bu ayrılma, birbirleriyle çiftleşip üreyemeyen canlı organizmalar oluşturabilir.[185]
İkinci türleşme mekanizması, canlıların küçük popülasyonlarının yeni bir çevrede izole olduklarında oluşan peripatrik türleşme şeklidir. Bu tür bir türleşme şekli, yalıtılan ve izole edilen popülasyonların sayısal olarak ebeveyn türden daha az olması ile allopatrik türleşme şeklinden ayrılır. Burada, kurucu etkisi, hızlı bir genetik değişime yol açan ve homozigot seçilimini artıran yakın akraba eşleşme oranının artması sonucu, hızlı bir türleşmeye yol açar.[186]
Üçüncü türleşme mekanizması, parapatrik türleşme şeklidir. Bu, küçük bir popülasyonun yeni bir yaşam ortamına ayak bastığı peripatrik türleşmeye benzer, ancak aradaki fark, bu iki popülasyon arasında fiziksel bir ayrılığın mevcut olmayışıdır. Bunun yerine, türleşme, daha ziyade popülasyonlar arasındaki gen akışını azaltarak indiren evrim mekanizmaları sonucu oluşur.[187] Bu, genellikle, ebeveyn türün yaşam alanları içinde ciddi çevre değişiklikleri olduğunda gerçekleşir. Buna örnek, maden ocakları çevresinde tespit edilen metal kirliliğine tepki olarak parapatrik türleşmeye uğrayan Anthoxanthum odoratum bitkisidir. Burada, bitkiler, topraktaki yüksek metal seviyesine karşı direnç geliştirmişlerdir. Metale karşı duyarlı ebeveyn popülasyonla yakın akraba eşleşmesine karşı seçilim, metale karşı dirençli olan bitkilerin çiçek açma dönemlerinde, sonunda tümüyle üretim yalıtımına yol açan kademeli bir değişime yol açar. Her iki popülasyon arasında oluşan hibritlere karşı seçilim, iki türün dış görünüşlerinde birbirlerinden giderek farklılaşmasında gözlemlenen karakterlerin yer değişimi gibi, bir tür içinde üremeyi destekleyen özelliklerin evrimini pekiştirmeye yol açabilir.[188]
Son olarak simpatrik türleşmede, türler, coğrafi yalıtım veya yaşadıkları habitatlarda çevresel değişim olmadan farklılaşırlar. Bu tür bir türleşme şekli, çok küçük miktardaki gen akışının, ayrı popülasyonlar arasındaki genetik farklılıkları ortadan kaldırdığından çok nadir gerçekleşir.[189] Genel olarak, hayvan türlerindeki simpatrik türleşme, üreme yalıtımının oluşmasına izin vererek hem genetik farklılıkların hem de rastlantısal olmayan eşleşmelerin evrimini teşvik eder.[190]
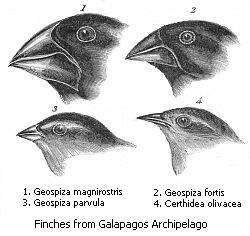
Sempatrik türleşmenin başka bir şekli olan poliploidizasyon yoluyla türleşme, birbirleriyle akraba olan iki türün, yeni bir hibrit tür oluşturduğu çapraz çiftleşimini içerir. Melez canlılar genelde kısır oldukları için, bu, hayvanlar aleminde yaygın olarak görülmez. Bunun nedeni, mayoz bölünme sırasında her bir ebeveynden gelen homolog kromozomların, aynı zamanda farklı türlerden gelmesi ve birbirleriyle başarılı olarak eşleşememesine dayanır. Ancak, bitkiler, poliploid oluşturmak için çoğunlukla kromozomlarını iki katına çıkardıklarından poliploidizasyon yoluyla türleşme, bitkiler aleminde daha yaygındır.[191] Bu da, her ebeveyn türden gelen kromozomların, her ebeveyn kromozomun hali hazırda zaten bir çift tarafından temsil edilmeleri dolayısıyla, mayoz bölünme sırasında kromozom çiftlerinin eşleşmesini şekillendirmesinde izin verir.[192] Böyle bir türleşme olayına örnek, bitki türleri Arabidopsis thaliana ile Arabidopsis arenosa'nın çapraz eşleşme sonucu melez bir tür olan Arabidopsis suecica bitki türünü ortaya çıkarmalarıdır.[193] Bu olay, yaklaşık 20.000 yıl önce gerçekleşmiş olup [194] bu türleşme süreci laboratuvarda da tekrarlanarak bu süreçte yer alan genetik mekanizmaların araştırılabilmesine olanak vermiştir.[195] Gerçekten de, katlanan kromozomların yarısı, bunun ikiye katlanmadığı canlılarla olan eşleşmelerde eşsiz kalacaklarından tek bir tür içindeki kromozom katlanması, üreme yalıtımının yaygın bir nedeni olabilir.[196]
Türleşme olayları, türlerin nispeten değişmeden kaldığı, görece uzun durgunluk dönemleri ile serpiştirilmiş fosil kayıtlardaki kısa evrim patlamaların izlerini de açıklaması bakımından Sıçramalı evrim teorisinde önemlidir.Niles Eldredge and Stephen Jay Gould, 1972. [197] Bu teoriye göre, türleşme ve hızlı bir evrim, yeni yaşam ortamlarında ve küçük popülasyonlarda türleşme geçiren organizmalar üzerinde en çok etki eden doğal seçilim ve genetik sürüklenme ile birlikte birbirleriyle bağlantılıdır. Sonuç olarak, fosil kayıtlarındaki durgunluk dönemi, ebeveyn popülasyona karşılık gelmekte, buna karşı türleşen ve hızlı evrim geçiren organizmalar ise, küçük bir popülasyonda bulundukları ve coğrafi olarak sınırlı bir yaşam alanında yaşadıkları için nadiren fosil olarak korunabilmişlerdir.[115]


Soy tükenmesi
Soy tükenmesi, bütün bir türün neslinin tükenerek yok olmasıdır. Türler, türleşme yoluyla devamlı olarak olarak ortaya çıktıklarından ve yok oluşlarda sürekli bir şekilde ortadan kalktıklarından, yok oluşlar seyrek görülen nadir olaylar değildir.[198] Dünya üzerinde yaşamış olan neredeyse tüm hayvan ve bitki türlerinin soyu, günümüzde tükenmiş olup [199] soy tükenmesi, tüm türlerin nihai kaderi gibi görülmektedir.[200]
Bu ani yok oluşlar, bazen tek tük oranda seyretmiş olsa da, bu tür soy tükenmesi olayları, yaşam tarihi boyunca sürekli meydana gelmiştir.[201] Kuş olmayan dinozorların yok olduğu Kretase-Tersiyer yok oluşu, en çok bilinen yok oluş olayı olmasına rağmen türlerin % 96'sının yok olduğu Permiyen-Triyas yok oluşu bundan daha şiddetli ve daha yıkıcı olmuştur.[201]
Holosen yok oluşu, geçtiğimiz birkaç bin yıl içinde insanlığın dünya geneline yayılması ile ilişkilendirilen bir yok oluş olayı olup hâlâ devam etmektedir. Günümüzdeki yok oluş oranları, arka plandaki orandan 100 ile 1000 kat daha büyük olup 21. yüzyılın ortalarında türlerin % 30'unun nesli tükenmesi beklenmektedir.[202]
Günümüzde devam etmekte olan yok oluşun birincil ve en başta gelen nedeni, insan faaliyetleri olup küresel ısınma gelecekte de bu yok oluşu hızlandıracağa benziyor.[203]
Yok oluşların evrimsel rolü, henüz pek iyi anlaşılmamış olup ne tür bir yok oluşun farz edildiği ile ilişkili olabilir.[201] Yok oluşların büyük bir çoğunluğunu oluşturan aralıksız ve sürekli oluşan düşük seviyedeki yok oluşların nedeni, sınırlı kaynaklar yüzünden türler arasında oluşan rekabetler sonucu olabilir (Rekabetçi dışlanım veya Gause kuralı).[204] Eğer bir tür başka bir türü rekabet dışı bırakırsa bu durum, daha elverişli ve uygun olan türün hayatta kaldığı ve diğer türün yok olmasına neden olan tür seçilimini ortaya çıkarabilir.[83]
Periyotik aralıklarla oluşan bu yok oluşlar da önemlidir, ancak bu tür yok oluşlar, seçici bir güç gibi hareket etmek yerine, büyük ölçüde ve belirli olmayan bir şekilde tür çeşitliliğini azaltırlar ve hızlı bir evrim patlamasına (adaptif radyasyon) ve hayatta kalabilen türlerin türleşmesine yol açarlar.[205]
Yaşamın evrimsel tarihi
- Ayrıca bakınız: Evrim zaman çizelgesi ve İnsan evrimi zaman çizelgesi
Hayatın kökeni
Hayatın ilk kez ortaya çıkışı, biyolojik evrim için temel bir ön şarttır, ancak evrimin işleyişini anlamak için hayatın kökeninin bulunması gerekli değildir,[206] çünkü bir kez canlı organizmalar ortaya çıktığında evrim kurallarının işleyeceği deneylerle gözlenmiştir.[207][208][209][210][211] Evrim için ilk organizma sorunu henüz tam anlamıyla çözülememiştir. Ortaya çıkan ilk canlı organizma hakkında çeşitli teoriler bulunmaktadır.
Şu anki bilimsel konsensüs karmaşık biyokimyanın, basit kimyasal reaksiyonlar ile hayatı oluşturduğu yönündedir, ancak bunun nasıl olduğu henüz tam anlamıyla çözülememiştir.[212] Hayatın ilk kez ortaya çıkışı, yaşayan ilk şeylerin yapısı veya evrensel ortak atanın genetik yapısı ile ilgili bilgiler henüz eksiktir.[213][214] Dolayısıyla, hayatın tam olarak nasıl başladığı konusunda bir konsensüs bulunmamaktadır, ancak RNA gibi kendini kopyalayan moleküller[215] ve basit hücre yapıları[216] ile ilgili teoriler mevcuttur.
Ortak ata
- Daha fazla bilgi: Ortak ata, Ortak atanın kanıtı ve Homoloji (biyoloji)
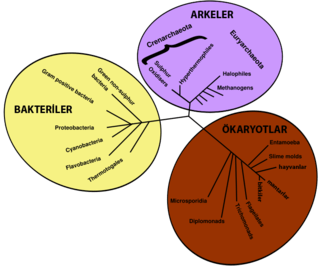
Yeryüzünde yaşayan tüm canlılar, ortak bir atadan veya bu ortak atanın gen havuzundan farklılaşarak türemiştir.[217][218] Günümüzdeki mevcut türler, uzun türleşme süreçlerinin bir sonucu olan çeşitlilikleri ve soy tükenmesi olayları ile evrim sürecinin çeşitli aşama ve evrelerini oluşturur.[219] Daha önce canlıların ortak bir atadan türedikleri sonucuna, canlıların sahip oldukları dört basit gerçekten yola çıkılarak varılmıştır: Birincisi, canlıların coğrafi dağılımları, her zaman yerel adaptasyonlarla açıklanamaz. İkincisi, canlı çeşitliliği, tamamen emsalsiz veya eşsiz olan, kendine mahsus canlılardan oluşmamaktadır, bilakis canlılar ortak morfolojik benzerlikler gösterirler. Üçüncü olarak, açık ve belirgin bir amacı olmayan körelmiş özellikler, daha önce türedikleri atalarının işlevsel özelliklerini gösterirler ve sonuncu olarak, canlılar, bu özellikler kullanılarak, bir aile ağacında olduğu gibi, bir hiyerarşi içinde içi içe yuvalanmış gruplar halinde sınıflandırılabilirler.[220] Ancak, modern araştırmalar, bazı genlerin, yatay gen transferi sonucu uzaktan akraba olan türler arasında da bağımsız olarak yayılabildiklerini gösterdiğinden, bu "evrimsel hayat ağacı", sandığımız şekilde basitçe dallara ayrılan bir ağaçtan daha çok karmaşık olabilir.[221][222]
Daha önce yaşamış türler de, evrimsel tarihlerine dair kayıt bulgular geride bırakmıştır. Fosiller, günümüz canlıların karşılaştırmalı anatomileri ile birlikte, canlılara dair morfolojik veya anatomik kayıtları oluşturur.[223] Modern ve soyu tükenmiş türlerin anatomilerinin karşılaştırılması yoluyla paleontologlar, bu türlerin soy ağacını ve sistematik sınıflandırılmalarını yaparlar. Ancak, bu yöntem, kabuk, kemik ve diş gibi, sert vücut veya uzuv parçalarına sahip olan canlılarda daha başarılı sonuçlar verir. Ayrıca, bakteri ve arkeler gibi prokaryotların fosilleri, ataları hakkında bilgi vermediği için, sınırlı sayıda morfolojik özellik dizileri gösterirler.
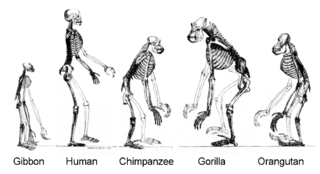
Yakın bir zaman önce, ortak ataya dair kanıtlar, canlılar arasındaki biyokimyasal benzerliklere dair yapılan bir araştırmada ortaya çıktı. Örneğin, tüm canlı hücreler, aynı temel nükleotit ve amino asit küme dizilerini kullanırlar.[224] Moleküler genetiğinde gelişmeler, canlı organizmaların genomunda, türlerin mutasyonların oluşturduğu moleküler saat tarafından ayrıldıkları zamanlardan kalma evrimsel kayıtların yer aldığını ortaya çıkarmıştır.[225] Örneğin, bu DNA dizisi karşılaştırmaları, insanlarla şempanzelerin, genomlarının % 96'sını paylaştığını ortaya koymuş ve ayrıldıkları birkaç bölgenin analizi ise, bu türlerin ortak atasının ne zaman yaşadığının belirlenmesine ışık tutmuştur.[226]
Canlıların evrimi
- Daha fazla bilgi: Evrim zaman çizelgesi
Prokaryotlar, yeryüzünde yaklaşık 3-4 milyar yıl önce yaşamış olup[227][228] morfolojik ve hücresel yapı organizasyonlarında gözle görülür belli bir değişiklik geçirmeden, önümüzdeki birkaç milyar yıl boyunca da yaşamaya devam ettiler.[229]
Ökaryotik hücreler, 1.6 ile 2.7 milyar yıl önce ortaya çıktılar. Hücre yapısındaki bir sonraki değişiklik, endosimbiyoz denilen bir simbiyoz ilişki içinde, ökaryot hücrelerin bakterileri yutması ve kendi bünyelerine almasıyla ortaya çıktı.[230][231] Yutulan bakteri ile konak hücre, daha sonra bakterinin mitokondriye ya da hidrogenozome dönüşmesiyle, birlikte evrim geçirdiler.[232] Siyanobakteri benzeri bakterilerin yutulduğu başka bir olay ise, bitki ve yosunlarda kloroplastların oluşumuna yol açtı.[233]
Yaşam tarihi, 610 milyon yıl öncesine kadar tek hücreli ökaryotlar, prokaryotlar ve arkeaların ardından çok hücreli organizmaların Edikara döneminde, okyanuslarda görünmeye başlamasıyla devam etmiştir.[227][234] Çok hücreliliğin evrimi, süngerler, kahverengi algler, siyanobakteriler, cıvık mantarlar ve miksobakteriler gibi canlılarda birden çok bağımsız olaylarla gelişmiştir.[235]
İlk çok hücreli canlıların ortaya çıkmasından kısa bir süre sonra, Kambriyen patlaması denilen bir olayda, yaklaşık 10 milyon yıl içinde, kayda değer miktarda bir biyolojik çeşitlilik ortaya çıktı. Bu dönemde, günümüzde yaşayan modern canlı türü tiplerinin büyük çoğunluğu fosil kayıtlarda görülmeye başladığı gibi sonradan soyları tükenen, benzersiz ve emsalleri olmayan canlı tür ve tiplerine rastlanır.[236] Kambriyen patlamasının tetikleyen nedenlere dair, fotosentez sonucu oluşan oksijenin atmosferde birikmesi veya Oksijen Felaketi gibi, çeşitli etkenler öne sürülmüştür.[237]
Yaklaşık 500 milyon yıl önce, ilk olarak bitkiler ve mantarlar, karalara yerleşmeye başladı ve hemen ardından bu süreci, eklem bacaklılar ile diğer hayvanların karaya çıkması takip etti.[238] Böcekler, özellikle başarılı bir tür olup günümüzde de hayvan türlerinin çoğunluğunu oluştururlar.[239] İki yaşamlılar (amfibiler), yaklaşık 364 milyon yıl önce ilk defa görülmeye başlarlar, ardından erken dönem amniyotları, sonra yaklaşık 155 milyon yıl önce kuşlar ile sürüngen benzeri türler ve 129 milyon yıl önce de memeliler takip eder.[240][241][242] Ancak, büyük ve gelişmiş yapılı bu hayvanların evrimine rağmen, daha erken dönemlerde ortaya çıkan küçük organizmalar gibi canlılar, hâlâ olsun çok başarılı türler olup biyokütlenin ve prokaryot türlerin büyük çoğunluğunu oluşturarak yeryüzüne hakimdirler.[243]
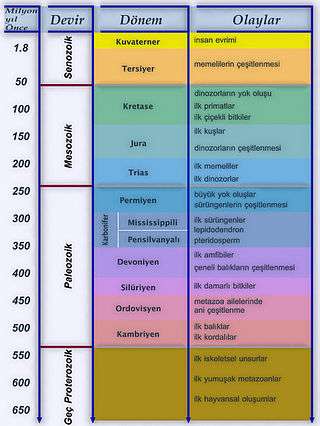
Aşağıdaki zaman çizelgesi, canlıların önemli evrimsel gelişmelerine dair genel bir bakış sunmaktadır.
| Jeolojik dönemler | Milyon yıl önce[244] | Gelişmeler |
|---|---|---|
| Kuvaterner | 2,588 | Buzul Çağına özgü hayvan ve bitki türleri |
| Paleojen ve Neojen (Tersiyer) | 65,5 | Memelilerin yayılımı; ilk primatlar ve büyük insansı maymunlar |
| Kretase | 145,5 | Kapalı tohumluların evrimi ve dinozorların da dahil olduğu kitlesel yok oluşların görülmesi |
| Jura | 199,6 | Dinozorların yayılımı, ilk kuşlar (Archaeopteryx) |
| Trias | 251 | İlk memeli canlılar, dinozorlar ve uçan sürüngenler |
| Permiyen | 299 | Sürüngenlerin yayılımı, „Memeli benzeri canlılar“, kitlesel yok oluş |
| Karbonifer | 359,2 | İki yaşamlıların (amfibiler) yayılımı, ilk kanatlı böcekler[245] |
| Devoniyen | 416 | Eğrelti otu, atkuyruğu ve kurtayağı (Lycopodium) bitkilerinin evrimi, omurgalıların sudan karaya çıkışı[246] |
| Silüryen | 443,7 | İlk damarlı bitkiler[247], eklem bacaklıların sudan karaya çıkışı[248], balık türlerin yayılımı |
| Ordovisyen | 488,3 | İlk kara bitkilerinin görünüşü[249], ilk "çeneli balıklar" |
| Kambriyen | 542 | „Burgess Şeyl faunası“, eklem bacaklılar, kordalılar (ve konodontlar), omurgalılar (çenesiz balıklar)[250] ve kafadan bacaklıların[251] görülmeye başlaması |
| Proterozoik (Erken dönem Dünya) | 2.5 milyar yıl önce | Mantarlar, ilk çok hücreli hayvanlar: süngerler[252], sölenterler, çift simetrili hayvanlar ve "Edikara faunası" |
| Arkeyan Devir (İlk dönem Dünya) | 4.0 milyar yıl önce | Tek hücreli yaşamın başlangıcı (Bakteriler, Arkeler, Ökaryotlar), Stromatolitler |
Bilimsel statüsü
Modern bilimde kuram, tutarlı bir bütün oluşturan gerçekler ve açıklamalardır. Modern fiziğin temel taşlarından olan Görelilik ve Kuantum kuramları, şu an üzerinde deliller toplanan, yeteri kadar test edilip güven verdiklerinde kanun konumuna yükselecek hipotezler değillerdir. Zaten bu olsa bile modern bilimde hiçbir teori bilimin sürekli değişebilirlik ilkesine göre kanun statüsüne konulamaz.[253] Evrim kuramı da aynı statüye sahiptir. Biyolojideki birçok veriyi birleştirip anlaşılır kılar; henüz kanıtlanmamış, test aşamasında olan bir "tahmin" değildir.
Evrim kuramı, insanlığın kökenine ilişkin sonuçları nedeniyle ortaya atıldığından bu yana sosyal ve politik alanda en çok tartışılan bilimsel kuramdır. Bunun sonucunda, kuramın bilimsel algılanışı ile popüler algılanışı oldukça farklı olagelmiştir. Evrim kuramına popüler düzeyde karşı çıkan ve onun yerine yeryüzündeki canlılığın kökeni ve çeşitliliğini doğaüstü bir yaratıcıya bağlayan akımlara genel olarak yaratılışçılık adı verilir.
Evrim kuramı, üç hususta açıklamalar getirir:[1]
- Evrimin olgusu (İng: fact of evolution) - canlı organizmaların ortak atalardan geldikleri ve birbirleri ile akraba oldukları bilgisi.
- Evrimsel tarih - Türlerin birbirlerinden tam olarak ne zaman ayrıldıkları ve bu ayrımların detayları.
- Evrimi gerçekleştiren mekanizma ve süreçler.
Bu basamaklardan birincisi olan evrimin olgusu, evrimin temel taşı ve son derece kesinlik arzeden bilgilere sahip olunan kısmıdır. Bu hususta Darwin'in topladığı birçok delilin üzerine yüzyıllardır birçok farklı biyoloji dalı tarafından toplanan deliller eklenmiştir. Günümüzde organizmaların evrimsel kökenlerine dair sahip olunan bilgiler, dünyanın yuvarlaklığı, gezegenlerin hareketleri ya da maddenin moleküler yapısı kadar "kesinlik arzeden" bilimsel çıkarımlardır.[1] Burada kastedilen kesinlik, şüphe götürmez bir gerçekliği ifade etmektedir. Diğer iki husustaki bilimsel çalışmalar ise aralıksız devam etmekte, her geçen gün yeni bir sonuca ulaşılmaktadır. Örneğin şempanze ve gorilin insana olan yakınlığının, babun veya diğer maymunlara olan yakınlıklarından daha fazla olduğu bugün kesin olarak bilinmektedir.[1]
Evrim kuramının bilimsel statüsü, eğitim, din, felsefe, bilim ve politika bağlamında sıkça gündeme getirilmektedir. Bu konu daha çok Amerika Birleşik Devletleri'nde Hristiyan cemaat ve lobilerin öncülüğünde gündeme gelmektedir. Fakat diğer ülkelerde, eğitim ve politikaya uzanmaya çalışan yaratılışçı görüşlerin savunucuları tarafından da gündeme getirilmektedir. Evrim kuramını destekleyen reddedilemez kanıtlar ve neredeyse mutlak denebilecek derecede bir bilimsel konsensüs olmasına rağmen, yaratılışçı şeklinde adlandırılan çevrelerce bilim dünyasında iki kutup varmış gibi gösterilmeye çalışılır. Yaratılışçı çevreler Amerika Birleşik Devletleri'nde, toplumdan büyük oranda destek görmediği iddiası ile Evrim Kuramı'nın okullarda bilim derslerinde okutulmasına karşı çıkmaktadır. Bu konuda Amerika'da yüzbinlerce bilim insanını temsil eden bilimsel meslek kurumları ve onun yanında 72 Nobel ödülü sahibi bilim insanı Evrim Kuramı'nı destekleyen bildiriler yayınlamıştır. Buna ek olarak açılan davalarda evrim kuramının bilimsel olduğu kabul görmüş bir teori olarak kabul edilmiş ve okullarda okutulmasının devamına karar verilmiştir.
Bilimsel camianın büyük bölümü, biyoloji, paleontoloji, antropoloji ve diğer disiplinlerdeki görüngüleri açıklayan yegane kuramın Evrim Kuramı olduğunda hemfikirdir. 1987 de yapılan bir araştırmanın sonuçlarına göre Amerika'daki doğa bilimleri alanında çalışan 500,000 bilim insanından yaklaşık %99.85'lik bir bölümünün evrim teorisini desteklediği ortaya konulmuştur. Evrim-yaratılış tartışmalarında uzman konumunda olan Brian Alters, doğa bilimleri alanlarında çalışan tüm bilim insanlarının %99.9'unun Evrim Kuramı'nı desteklediğini belirtmiştir. Benzer şekilde, dünyanın değişik ülkelerindeki bilimsel çevreler defalarca Evrim Kuramı'nın bilimsel olduğuna ilişkin bildiriler yayınlamıştır.[254][255][256][257][258] 1987 yılında Amerika'daki bilim insanları arasında yapılan bir araştırma, 480.000 bilim insanından sadece 700 bilim insanının yaratılışçı ve benzeri açıklamalara itibar ettiğini ya da Evrim Kuramı'na karşı şüphe duyduğunu göstermiştir. Ve bu 700 (%0.158) bilim insanından sadece küçük bir bölümü doğa bilimleri alanında akademik çalışma yapmaktadır.[259] Son yıllarda yapılan benzeri karşılaştırmalar, Evrim Kuramı'nı bütünü ile reddeden ya da ona karşı şüphe duyan bilim insanlarının oranının yaklaşık olarak %0.054 civarında olduğunu göstermiştir. Karşı çıkanların %75.1'i biyoloji dışındaki bilim dallarında çalışmaktadır.
Son yıllarda genetik biliminin gelişmesi ile kuş embriyoları üzerinde çalışan bilim insanları bazı kapalı genleri açarak kuş embriyolarında kuşların evrimsel atası olan dinozor embriyolarındakine benzer görüntülere (diş, pul, boyun ve kuyruk yapısı) ulaşmışlar,[260][261] Avustralya'daki diğer bilim insanları ise değişik canlılar üzerinde evrimin hızını (yüz bin yılda gerçekleşen genetik mutasyon oranı) ölçmeyi başarmışlardır.[262][263]
Steve Projesi
Yaratılışçıların, "evrim konusunda bilimsel konsensüs olmadığı" yönündeki iddialarını çürütmek için, ABD Ulusal Bilimler Akademisi, Steve Projesi'ni başlatmıştır. Bu projenin amacı, isminde sadece Steve geçen bilim insanlarının kaç tanesinin Evrim Kuramı'nı desteklediğini ortaya koymaktır. Ortaya çıkan liste (Steve-o-metre) çoğunluğu biyoloji dallarında çalışan, isimlerinde Steve sözcüğü ya da bu ismin değişik telaffuzları geçen bilim insanlarını sıralamakta ve Yaratılışçılar ile Yeni Yaratılışçıların yayınlamış oldukları listelerden daha kalabalık olduğunu göstermektedir.
Evrimi kabul eden sadece Steve isimli bilimadamları, evrimi kabul etmeyen tüm bilim insanlarından daha fazladır. Bu projede James gibi çok daha yaygın (1. sırada[264]) bir isim yerine Steve gibi çok daha az kullanılan (74. sırada[265]) bir ismin seçilmesi de araştırmanın sonuçlarının güvenilirliğini desteklemektedir.
Bilimsel konular, elbette kimin listesinin daha uzun olduğu temelinde tartışılmamalıdır fakat dünyada bilim dünyasında bir çelişkinin olmadığını, tam tersine çok güçlü bir konsensüsün olduğunu göstermesi açısından Steve Projesi eğlendirici bir örnektir.
Uygulama alanları
- Daha fazla bilgi: Yapay seçilim ve Genetik algoritma
Evrimsel biyolojide kullanılan kavram ve modellerin, özellikle doğal seçilimde, birçok uygulamaları mevcuttur.[266]
Yapay seçilim, canlı popülasyondaki istenilir bir özelliğin bilinçli olarak seçilimidir. Yapay seçilim yöntemleri, hayvan ve bitkilerin evcilleştirilmesinde binlerce yıldır kullanılmaktadır.[267] Daha yakın bir zamanda, bu tür yapay seçilim yöntemleri, antibiyotik direnç genleri oluşturmada ve DNA yapısını değiştirmede seçilebilir işaretler kullanılarak genetik mühendisliğin önemli bir parçası haline gelmiştir. Laboratuvar ortamında oluşturulan mutasyon döngülerinde ve bunu izleyen yapay seçilimde, yönlendirilmiş seçilim denilen bir süreç ile, örneğin modifiye edilmiş enzimler ve yeni antikorlar gibi değerli özelliklere sahip proteinler evrilebilmiştir.[268]
Organizmanın evrimi sırasında meydana gelen değişiklikleri anlamak, yeni vücut parçaları ve uzuvları oluşturmak için gerekli olan genler ile insanlardaki genetik bozukluklar ile ilişkili olabilecek diğer genleri ortaya çıkarabilir.[269] Örneğin, Astyanax mexicanus evrimsel gelişimi sırasında görme özelliğini kaybetmiş olan ve karanlıkta yaşayan albino bir mağara balığıdır. Görme özelliğini yitirmiş bu kör balık türünün farklı popülasyonlarının birbirleriyle üremesi sonucu, farklı mağaralarda evrimleşmiş olan yalıtılmış popülasyonlarda farklı mutasyonlar ortaya çıktığı için görme özelliğine sahip yavru bireyler meydana gelebilmektedir.[270] Bu gözlem, görme ve pigmantasyon için gerekli olan genleri tespit etmede yardımcı olmuştur.[271]
Bilişim biliminde, evrimsel algoritmalar ve yapay yaşam kullanılarak oluşturulan evrim simülasyonları 1960'lı yıllarda başlamış ve yapay seçilim simülasyonları ile genişletilerek devam etmiştir.[272] Yapay evrim, İngo Rechenberg'in çalışmalarının bir sonucu olarak 1960'lı yıllarda yaygın olarak tanınan bir optimizasyon yöntemi oldu. Rechenberg, karmaşık mühendislik problemlerini çözmek için evrim stratejileri kullanmıştır.[273] Genetik algoritmalar, özellikle John Holland'ın çalışmaları sayesinde popülerlik kazanmıştır.[274] Pratik uygulamalar arasında, bilgisayar programlarının özgüdümsel olarak kendi kendilerini geliştirmeleri ve otomatik olarak evrilmeleri de gösterilebilir.[275] Evrimsel algoritmalar günümüzde, tasarımcılar yani insanlar tarafından üretilen yazılımlardan daha verimli bir şekilde çok boyutlu sorunları çözmek ve aynı zamanda sistemlerin tasarımını optize etmek için kullanılmaktadır.[276]
Sosyal ve kültürel tepkiler
- Ayrıca bakınız: Akıllı tasarım davası ve Scopes Davası
.jpg)
19. yüzyılda, özellikle 1859 yılında "Türlerin Kökeni" isimli kitabın yayınlanmasından sonra, yaşamın evrimleştiği fikri, evrimin felsefi, sosyal, dini çıkarımları hakkındaki akademik tartışmaların önemli bir kaynağı olmuştur. Günümüzde, modern evrimsel sentez, bilim insanlarının büyük bir çoğunluğu tarafından kabul edilmektedir.[278] Ancak, evrim, bazı teistler için tartışmalı bir kavram olarak kalmıştır.[279]
Teistik evrim gibi kavramlar üzerinden kendi inançları ile evrim olgusu arasında bir uzlaşı sağlayabilmiş çeşitli din ve mezhepler olduğu gibi, evrimin dinsel kaynaklarda yer alan yaratılış mitleriyle çeliştiğine inanan ve evrim olgusuna karşı itirazlarda bulunan yaratılışçılar da bulunmaktadır.[280][281][282] 1844 yılında yayınlanan "Doğal Yaratılış Tarihinin İzleri" (İng:Vestiges of the Natural History of Creation) isimli bir kitapta verilen tepkilerin gösterdiği gibi, evrimsel biyolojinin en tartışmalı yönünün, insanların genel olarak maymun olarak adlandırılan şempanze, goril gibi canlılarla ortak bir ataya sahip olması ve modern taksonomide insanın da bir maymun türü olarak kabul ediliyor olması insanlığın zihinsel ve ahlâki birimlerinin, hayvanlardaki diğer kalıtımsal özellikler gibi doğal nedenlere dayandığı ve genelde insan evrimi ile ilgili olan diğer çıkarımlar olduğu görülmektedir.[283] Bazı ülkelerde, özellikle Amerika Birleşik Devletleri'nde, bilim ve din arasındaki bu gerilimler, siyasete ve millî eğitime odaklanmış dinsel bir çatışma şeklinde günümüzde mevcut olan Yaratılışçılık ve Evrim Kuramı tartışmalarının ortaya çıkmasına yol açmıştır.[284] Kozmoloji ve yer bilimleri gibi diğer bilimsel alanlarda da birçok dini metinlerin yorumlanması ile ilgili tartışmalar mevcut olsa da, evrimsel biyoloji bunlardan çok daha fazla bir dini muhalefete maruz kalmaktadır.
Evrimin Amerikan lise biyoloji derslerindeki öğretimi, 20. yüzyılın ilk yarısında fazlaca yaygın değildi. Maymun Davası olarak da bilinen 1925 tarihli Scopes Davası'nda alınan kararlar, evrim konusunun Amerikan lise biyoloji kitaplarında genç nesiller için çok nadir olarak ele alınıp işlenmesine ve evrim karşıtı yasaların 1960'lı yıllara kadar çeşitli eyaletlerde yürürlükte kalmasına neden olmuştur. Ancak bir nesil sonra, evrim konusu biyoloji derslerinde yavaş yavaş tekrar ele alınmaya başlanmış ve 1968 tarihinde bir devlet lisesinde biyoloji öğretmeni olan Susan Epperson'un bu durumu mahkemeye götürmesi ile sonuçlanan Epperson vs. Arkansas davasının temyiz sonuçları ardından, bilimsel bir teorinin yasaklanmasının anayasaya aykırı olduğu kararı alınarak evrim kuramı yasal olarak korunma altına alınmıştır. O zamandan beri, yaratılışçıların eğitim müfredatındaki hâkimiyetleri azalmaya başlayarak bunu takip eden 1970'li ve 1980'li yıllarda, eğitim programlarında alınan çeşitli kararlarla, orta öğretim müfredatında karşıt yaratılışçı düşüncelerin öğretilmesine yasal olarak izin verilmemiş ve bu sefer Akıllı tasarım olarak geri döndüğünde, Dover davası olarak da bilinen 2005 tarihli Kitzmiller v. Dover Area School davasında akıllı tasarımın bilim olmadığı ve okullarda bilimmiş gibi öğretilemeyeceğine karar verilerek bir kez daha devlet okullarından dışlanmıştır.[285]
Günümüzde halkın evrime bakışı
| Dini inanca göre ABD'de evrime destek oranının dağılımı | |||||||||||
|
Budist
81 |
Hindu
80 |
Musevi
77 |
Bağlımsız
72 |
Katolik
58 |
Ortodoks
54 |
Protestan
51 |
Müslüman
45 |
Siyah Prot.
38 |
Evanjelik
24 |
Mormon
22 |
Yehova Şahitleri
8 |
| Kaynak:U.S.Religious Landscape Survey (6/2008)[286] Açıklama:Evrimin yeryüzünde insanın kökenine dair en iyi açıklama olduğunu kabul edenlerin oranı % | |||||||||||
2005 yılında, ABD Michigan Eyalet Üniversitesi’nden bir grup bilim insanının hazırladığı ve gelişmiş ülkeler seviyesindeki 34 ülkeyi içeren bir çalışmada, "evrimi doğru kabul edenlerin oranı" yaklaşık %27 ile en düşük Türkiye'de bulunmuştur.[287] Türkiye'den sonra ise %40 ile, akıllı tasarım akımının ortaya çıktığı ABD yer almaktadır. Yine aynı araştırmaya göre, özellikle insanın evrimi en fazla Türkiye’de % 51 ile reddedilmişken ardından Türkiye’yi % 39 ile ABD izlemektedir.
Gelişmiş Avrupa devletlerinde evrimin doğru kabul edilme oranları Türkiye'den ve ABD'dan çok daha yüksektir. İzlanda'da halkın %80'inden fazlası, Danimarka, Fransa, Birleşik Krallık, Japonya'da yaklaşık %80'i evrimi kesin olarak doğru kabul etmektedir. Geri kalanların büyük bir kısmı ise emin olmadığını belirtmiştir.[287] ABD Michigan Eyalet Üniversitesi’nden Prof. Jon D. Miller ve ekibinin yayınladıkları araştırmaya göre örneğin, ABD'de 1484 denek arasında “bugün bildiğimiz insan daha önceki hayvan türlerinden evrimleşerek gelişmiştir” ifadesini doğru bulanların oranı % 40 iken, ifadeyi "yanlış" bulanların oranı % 39 ve "emin olmayanların" oranı ise % 21 olarak belirtilmiştir. Türkiye’deki oran ise sırasıyla, "doğru" % 27, "yanlış" % 22 ve "emin değilim" % 51 olarak verilmiştir. Soru, farklı bir şekilde “insanlar Tanrı tarafından bugünkü haliyle yaratıldılar, yaşamın daha önceki biçimlerinden evrilmediler” diye sorulduğunda, evrimi reddedenlerin oranı % 62 olarak artmış, "yanlış" bulanların oranı % 36 iken "emin olmayanların" oranı ise % 2'de kalmıştır.[288]
ABD’li deneklerin insanın evrimine dair tereddütleri ve itirazları varken buna karşın hayvan ve bitkilerin evrimini ise daha kolay kabullendikleri de bu araştırmanın ortaya çıkardığı başka bir sonuçtur. Nitekim soru değiştirildiğinde, “bazı bitki ve hayvan türleri, milyonlarca yıllık süre içinde uyum sağlayıp hayatta kalırken, diğer türler öldüler ve nesilleri tükendi” ifadesini "doğru" bulanların oranı % 78 ile yüksek olup "yanlış" bulanların oranı ise sadece % 6 oranında seyretmektedir.[289]
Miller, Scott ve Okamoto’nun (2005 ve 2006) 34 ülkede yaptıkları bu çalışmanın sonuçları evrimsel yaklaşıma ilişkin gittikçe azalan bir genel kabul düzeyine işaret etmektedir. Nitekim, bu çalışmanın önemli bulgularından birisi de, Türkiye ve ABD’de, evrimsel süreçten habersiz ya da evrim konusunda kararsız olanların oranı 1985 yılında %7 iken, bu oranın 2005 yılında %21’e yükselmiş olmasıdır. Böylece, bu ülkelerde her beş yetişkinden birisi evrimden ya habersiz ya da bu konuda kararsız hale gelmiştir.[290]
Türkiye
Bilim ve Ütopya dergisinde Ekim 2001 tarihinde yayınlanan "Safsata Anketi" isimli başka bir araştırmada ise İstanbul Üniversitesi, Ankara Üniversitesi ve ODTÜ gibi çeşitli üniversitelerin tıp, biyoloji, fizik, kimya, astronomi, jeoloji gibi doğa bilimleri fakültelerindeki 1. ve 4. sınıf öğrencilerine "insan soyunun Havva ve Adem’den geldiği görüşüne inanıp inanmadıkları" sorulmuş ve İÜ Fen Fakültesi Biyoloji Bölümü 1. sınıf öğrencileri bu soruya % 73.13 ile "inanıyorum", % 16.42 ile "olabilir" ve % 10.45 ile "inanmıyorum" cevabı vermiştir. Aynı üniversitenin 4. sınıf Biyoloji bölümü öğrencileri ise % 75.00 "inanıyorum", % 18.75 "olabilir" ve % 6.25 "inanmıyorum" olarak yanıtlamışlardır. Bu oran İÜ Cerrahpaşa Tıp Fakültesi 1. sınıf öğrencilerinde sırasına göre % 66.67, % 16.67 ve % 16.67 iken 4. sınıf öğrencilerinde ise % 74.71, % 9.20 ve % 16.09 ve İÜ İstanbul Tıp Fakültesi 1. sınıf öğrencilerinde % 67.86, % 21.43 ve % 10.71 iken, 4. sınıf öğrencilerinde ise % 80.00, % 20.00 ve % 0.00 olmuştur.[291]
Halkın evrime bakış açısının altında yatan nedenlere dair Türkiye'nin kendine özgü sorunları olduğu (Dr. Kenan Ateş), Türkiye’nin hızlı küreselleşmeye hazırlıksız yakalanmış olması (Seçkin ve Okçabol, 2006), Kur’an’da anlatılan insanın yaratılış öyküsününün bilimin bulgularıyla çeliştiğinin düşünülmesi, bu anlamda insanı diğer canlılardan ayrı tutma eğilimi ve Türkiye'nin Avrupa'da olduğu gibi bir Aydınlanma döneminden geçmemiş olması gibi düşünceler öne sürülmüşse de evrim kuramının neden az benimsendiğine ve toplumdaki kabullenebilirlik seviyesine dair daha birçok sosyokültürel araştırmaların yapılması önem taşımaktadır.[289]
Brezilya
Brezilya'da yapılan 2010 tarihli bir ankette, teistik evrime inandıklarını söyleyen veya evrimin Tanrı tarafından yönlendirildiğini düşünenlerin oranı % 59 olarak tespit edilirken evrimi reddeden yaratılışçıların oranı % 25 ve evrimin herhangi bir ilahi güce gerek kalmadan oluştuğunu söyleyenlerin oranı ise % 8 olarak tespit edilmiştir.[292]
Kanada
Kanada'da yapılan bir araştırmada ise, insanın daha az gelişmiş yaşam formlarından oluştuğunu düşünenlerin oranı % 59 olarak tespit edilirken insan ve dinozorların yeryüzünde aynı zamanda ve aynı dönemde yaşadıklarına inanan Kanadalıların oranı ise % 42 olarak belirlenmiştir. Tanrının insanı son 10.000 yıl içinde bugünkü haliyle yarattığına inanan Kanadalıların oranı ise % 22.[293]
Birleşik Krallık
"Hayatın kökeni ve gelişimi" üzerine Birleşik Krallık'ta yapılan 2006 tarihli bir araştırma anketinde katılımcıların yaşamın kökenine dair üç farklı açıklama arasında bir seçim yapılmaları istenmiş ve deneklerin % 22'si Genç Dünya yaratılışçılığını, % 17'si ise Akıllı tasarımı ve % 48'i ise evrim kuramını (açıkça tanrısal bir rol olmadan) tercih etmişlerdir. Diğerleri ise kararsız kalmıştır.[294][295]
Avustralya
Avustralya'daki 2009 tarihli anket, Avustralyalıların neredeyse dörtte birinin insanın İncil'deki yaratılışına inandığını, % 32'nin Tanrı tarafından yönlendirilen bir evrime ve % 42'nin de yaşamın kökeni için "tamamen bilimsel" bir açıklamaya inandıklarını ortaya koymuştur.[296]
2010 yılında Auspoll ve Avustralya Bilim Akademisi tarafından yapılan bir anket ise Avustralyalıların yaklaşık % 80'inin, evrimin günümüzde de hâlâ oluştuğunu ve devam ettiğini düşündüklerini ortaya koymuştur. Bu ankette Avustralyalıların % 10'u evrime inanmazken % 11'i ise emin olmadıklarını belirtmiştir.[297]
Ayrıca bakınız
Kaynakça
- "Evrim." Encyclopædia Britannica. Encyclopædia Britannica Online. Encyclopædia Britannica Inc.
- Boğa boynuzlu akasya KillerPlants.com (İngilizce)
Dipnotlar
- 1 2 3 4 5 6 7 8 9 10 11 12 13 "evolution." Encyclopædia Britannica. Encyclopædia Britannica Online. Encyclopædia Britannica Inc. 2011
- ↑ Understanding Evolution: Mutations URL Erişim Tarihi: Ekim 2011
- 1 2 3 Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-187-2.
- ↑ Evrimi Anlamak
- ↑ Practical uses of evolution talkorigins.org. Erişim: 9 Kasım 2011. Arşiv: <http://www.webcitation.org/634aPyHba>
- 1 2 3 4 Relevance of evolutionary biology to the national research agenda.(present and future importance of evolutionary biology), BioScience | November 1, 1999, Prepared by delegates representing the following scientific societies,
- American Society of Naturalists
- Animal Behavior Society
- Ecological Society of America
- Genetics Society of America
- Paleontological Society
- Society for Molecular Biology and Evolution
- Society of Systematic Biologists
- Society for the Study of Evolution
- ↑ "Nothing in biology makes sense, except in the light of evolution!"
- ↑ The Early Superstitions of Medicine, The Popular Science Monthly, May 1872, Volume 1, pp95-100,
- ↑ http://www.evrimianlamak.org/e/P1:evrim_tarihi_2_3
- 1 2 http://www.evrimteorisi.org/index.php?option=com_content&view=article&id=283&Itemid=108
- ↑ http://www.evrimianlamak.org/e/P1:evrim_tarihi_2_4
- ↑ http://www.evrimianlamak.org/e/P1:evrim_tarihi_2_5
- ↑ Korotayev A., Malkov A., Khaltourina D. Introduction to Social Macrodynamics: Secular Cycles and Millennial Trends. Moscow: URSS, 2006. ISBN 5-484-00559-0
- ↑ Prof.Dr.Ali Demirsoy "Yaşamın Temel Kuralları", Meteksan Yayınları, Ankara 1990. ISBN 97577460307
- ↑ “1oo Prüfungsfragen Abitur Biologie” - Prof. Dr. Wilfried Probst, Bibliografisches İnstitut AG, Mannheim 2009
- ↑ Evolution Mensch” - Rosemarie Benke-Bursian, Gondram Yayınları, Bindlach 2006
- ↑ http://www.evrimianlamak.org/e/P1:evrim_tarihi_3_2
- ↑ “Evolution und Biologische Vielfalt” - Prof. Dr. Wilfried Probst, Bibliografisches İnstitut & F.A. Brockhaus, Mannheim 2008
- ↑ Bowler, Peter J. (2003). Evolution: the history of an idea. Berkeley: University of California Press. ISBN 0-520-23693-9.
- ↑ Hald, Anders (1998). A History of Mathematical Statistics. New York: Wiley. ISBN 0-471-17912-4.
- ↑ Ford, E. B. (November 1977). "Theodosius Grigorievich Dobzhansky, 25 January 1900 -- 18 December 1975". Biographical Memoirs of Fellows of the Royal Society 23: 58–89. DOI:10.1098/rsbm.1977.0004.
- ↑ Mayr, Ernst: Systematics and the Origin of Species from a Viewpoint of a Zoologist, Harvard University Press 1942
- ↑ Huxley, Julian (2010): Evolution - The Modern Synthesis, MIT Press
- ↑ Sturm RA, Frudakis TN (2004). "Eye colour: portals into pigmentation genes and ancestry". Trends Genet. 20 (8): 327–32. DOI:10.1016/j.tig.2004.06.010. PMID 15262401.
- 1 2 Pearson H (2006). "Genetics: what is a gene?". Nature 441 (7092): 398–401. Bibcode 2006Natur.441..398P. DOI:10.1038/441398a. PMID 16724031.
- ↑ Visscher PM, Hill WG, Wray NR (2008). "Heritability in the genomics era—concepts and misconceptions". Nat. Rev. Genet. 9 (4): 255–66. DOI:10.1038/nrg2322. PMID 18319743.
- ↑ Oetting WS, Brilliant MH, King RA (1996). "The clinical spectrum of albinism in humans". Molecular medicine today 2 (8): 330–5. DOI:10.1016/1357-4310(96)81798-9. PMID 8796918.
- ↑ Phillips PC (2008). "Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems". Nat. Rev. Genet. 9 (11): 855–67. DOI:10.1038/nrg2452. PMC 2689140. PMID 18852697. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2689140.
- ↑ Wu R, Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. 7 (3): 229–37. DOI:10.1038/nrg1804. PMID 16485021.
- ↑ Jablonka, E.; Raz, G. (2009). "Transgenerational epigenetic inheritance: Prevalence, mechanisms and implications for the study of heredity and evolution.". The Quarterly Review of Biology 84 (2): 131–176. DOI:10.1086/598822. PMID 19606595. http://compgen.unc.edu/wiki/images/d/df/JablonkaQtrRevBio2009.pdf.
- ↑ Bossdorf, O.; Arcuri, D.; Richards, C. L.; Pigliucci, M. (2010). "Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana". Evolutionary Ecology 24 (3): 541–553. DOI:10.1007/s10682-010-9372-7. http://www.springerlink.com/content/c847255ur67w2487/.
- ↑ Jablonka, E.; Lamb, M. (2005). Evolution in four dimensions: Genetic, epigenetic, behavioural and symbolic. MIT Press. ISBN 0-262-10107-6. http://books.google.ca/books?id=EaCiHFq3MWsC&printsec=frontcover.
- ↑ Jablonka, E.; Lamb, M. J. (2002). "The changing concept of epigenetics". Annals of the New York Academy of Sciences 981 (1): 82–96. Bibcode 2002NYASA.981...82J. DOI:10.1111/j.1749-6632.2002.tb04913.x. http://a-c-elitzur.co.il/uploads/articlesdocs/Jablonka.pdf.
- ↑ Laland, K. N.; Sterelny, K. (2006). "Perspective: Seven reasons (not) to neglect niche construction". Evolution 60 (8): 1751–1762. DOI:10.1111/j.0014-3820.2006.tb00520.x. http://lalandlab.st-andrews.ac.uk/pdf/laland_Evolution_2006.pdf.
- ↑ Chapman, M. J.; Margulis, L. (1998). "Morphogenesis by symbiogenesis". International Microbiology 1 (4): 319–326. PMID 10943381. http://www.im.microbios.org/04december98/14%20Chapman.pdf.
- ↑ Wilson, D. S.; Wilson, E. O. (2007). "Rethinking the theoretical foundation of sociobiology". The Quarterly Review of Biology 82 (4). http://evolution.binghamton.edu/dswilson/wp-content/uploads/2010/01/Rethinking-sociobiology.pdf.
- ↑ Wu R, Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits".Nat. Rev. Genet. 7 (3): 229–37.
- 1 2 Ewens W.J. (2004). Mathematical Population Genetics (2nd Edition). Springer-Verlag, New York. ISBN 0-387-20191-2.
- ↑ Butlin RK, Tregenza T (1998). "Levels of genetic polymorphism: marker loci versus quantitative traits". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 187–98.
- ↑ Wetterbom A, Sevov M, Cavelier L, Bergström TF (2006). "Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution" .J. Mol. Evol. 63 (5): 682–90.
- ↑ Sawyer SA, Parsch J, Zhang Z, Hartl DL (2007). "Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila" Proc. Natl. Acad. Sci. U.S.A. 104 (16): 6504–10
- ↑ Hastings, P J; Lupski, JR; Rosenberg, SM; Ira, G (2009). "Mechanisms of change in gene copy number". Nature Reviews. Genetics 10 (8): 551–564
- ↑ Carroll SB, Grenier J, Weatherbee SD (2005). From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Second Edition. Oxford: Blackwell Publishing. ISBN 1-4051-1950-0.
- ↑ Harrison P, Gerstein M (2002). "Studying genomes through the aeons: protein families, pseudogenes and proteome evolution". J Mol Biol 318 (5): 1155–74.
- ↑ Bowmaker JK (1998). "Evolution of colour vision in vertebrates". Eye (London, England)12 (Pt 3b): 541–7.
- ↑ Gregory TR, Hebert PD (1999). "The modulation of DNA content: proximate causes and ultimate consequences". Genome Res. 9 (4): 317–24.
- ↑ Hurles M 2004). "Gene duplication: the genomic trade in spare parts". PLoS Biol. 2 (7): E206.
- ↑ Liu N, Okamura K, Tyler DM (2008). "The evolution and functional diversification of animal microRNA genes" Cell Res. 18 (10): 985–96
- ↑ Siepel A (2009). "Darwinian alchemy: Human genes from noncoding DNA". Genome Res. 19 (10): 1693–5
- ↑ Orengo CA, Thornton JM (2005). "Protein families and their evolution-a structural perspective". Annu. Rev. Biochem. 74 (1): 867–900.
- ↑ Long M, Betrán E, Thornton K, Wang W (2003). "The origin of new genes: glimpses from the young and old". Nat. Rev. Genet. 4 (11): 865–75.
- ↑ Wang M, Caetano-Anollés G (2009). "The evolutionary mechanics of domain organization in proteomes and the rise of modularity in the protein world". Structure 17 (1): 66–78.
- ↑ Weissman KJ, Müller R (2008). "Protein-protein interactions in multienzyme megasynthetases". Chembiochem 9 (6): 826–48
- ↑ Radding C (1982). "Homologous pairing and strand exchange in genetic recombination". Annu. Rev. Genet. 16 (1): 405–37
- ↑ Agrawal AF (2006). "Evolution of sex: why do organisms shuffle their genotypes?". Curr. Biol. 16 (17): R696.
- ↑ Peters AD, Otto SP (2003). "Liberating genetic variance through sex". Bioessays 25 (6): 533–7.
- ↑ Goddard MR, Godfray HC, Burt A (2005). "Sex increases the efficacy of natural selection in experimental yeast populations". Nature 434 (7033): 636–40.
- 1 2 Morjan C, Rieseberg L (2004). "How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles". Mol. Ecol. 13 (6): 1341–56.
- ↑ Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF (2003)."Lateral gene transfer and the origins of prokaryotic groups". Annu Rev Genet 37 (1): 283–328.
- ↑ Walsh T (2006). "Combinatorial genetic evolution of multiresistance". Curr. Opin. Microbiol. 9 (5): 476–82.
- ↑ Kondo N, Nikoh N, Ijichi N, Shimada M, Fukatsu T (2002). "Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect". Proc. Natl. Acad. Sci. U.S.A. 99(22): 14280–5.
- ↑ Sprague G (1991). "Genetic exchange between kingdoms". Curr. Opin. Genet. Dev. 1 (4): 530–3.
- ↑ Gladyshev EA, Meselson M, Arkhipova IR (2008). "Massive horizontal gene transfer in bdelloid rotifers". Science 320 (5880): 1210–3.
- ↑ Baldo A, McClure M (1 September 1999). "Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts". J. Virol. 73 (9): 7710–21
- ↑ River, M. C. and Lake, J. A. (2004). "The ring of life provides evidence for a genome fusion origin of eukaryotes". Nature 431 (9): 152–5.
- ↑ Lande R, Arnold SJ (1983). "The measurement of selection on correlated characters". Evolution 37: 1210–26}. DOI:10.2307/2408842.
- ↑ Ayala FJ (2007). "Darwin's greatest discovery: design without designer". Proc. Natl. Acad. Sci. U.S.A. 104 Suppl 1: 8567–73. DOI:10.1073/pnas.0701072104. PMID 17494753. http://www.pnas.org/cgi/content/full/104/suppl_1/8567.
- 1 2 Hurst LD (2009). "Fundamental concepts in genetics: genetics and the understanding of selection". Nat. Rev. Genet. 10 (2): 83–93
- ↑ Orr HA (2009)."Fitness and its role in evolutionary genetics". Nat. Rev. Genet. 10 (8): 531–9.
- ↑ Haldane J (1959). "The theory of natural selection today". Nature 183 (4663): 710–3.
- ↑ Orr HA (2009). "Fitness and its role in evolutionary genetics". Nat. Rev. Genet. 10 (8): 531–9.
- ↑ Lande R, Arnold SJ (1983). "The measurement of selection on correlated characters". Evolution37 (6): 1210–26.
- ↑ Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-187-2.
- ↑ Goldberg, Emma E; Igić, B (2008). "On phylogenetic tests of irreversible evolution". Evolution 62 (11): 2727–2741.
- ↑ Collin, Rachel; Miglietta, MP (2008). "Reversing opinions on Dollo's Law". Trends in Ecology & Evolution 23 (11): 602–609.
- ↑ Hoekstra H, Hoekstra J, Berrigan D, Vignieri S, Hoang A, Hill C, Beerli P, Kingsolver J (2001). "Strength and tempo of directional selection in the wild". Proc. Natl. Acad. Sci. U.S.A. 98 (16): 9157–60
- ↑ Hurst LD (2009). "Fundamental concepts in genetics: genetics and the understanding of selection". Nat. Rev. Genet. 10 (2): 83–93.
- ↑ Felsenstein (1 November 1979). "Excursions along the Interface between Disruptive and Stabilizing Selection". Genetics 93 (3): 773–95.
- ↑ Andersson M, Simmons L (2006). "Sexual selection and mate choice". Trends Ecol. Evol. (Amst.) 21 (6): 296–302.
- ↑ Kokko H, Brooks R, McNamara J, Houston A (2002). "The sexual selection continuum". Proc. Biol. Sci. 269 (1498): 1331–40.
- ↑ Hunt J, Brooks R, Jennions M, Smith M, Bentsen C, Bussière L (2004). "High-quality male field crickets invest heavily in sexual display but die young". Nature 432 (7020): 1024–7.
- ↑ Odum, EP (1971) Fundamentals of ecology, third edition, Saunders New York
- 1 2 Gould SJ (1998). "Gulliver's further travels: the necessity and difficulty of a hierarchical theory of selection". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 307–14.
- ↑ Gould SJ (1998). "Gulliver's further travels: the necessity and difficulty of a hierarchical theory of selection" Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353 (1366): 307–14.
- ↑ Mayr E (1997). "The objects of selection". Proc. Natl. Acad. Sci. U.S.A. 94 (6): 2091–4.
- ↑ Maynard Smith J (1998). "The units of selection". Novartis Found. Symp. 213: 203–11; discussion 211–7
- ↑ Hickey DA (1992). "Evolutionary dynamics of transposable elements in prokaryotes and eukaryotes". Genetica 86 (1–3): 269–74.
- ↑ Gould SJ, Lloyd EA (1999). "Individuality and adaptation across levels of selection: how shall we name and generalise the unit of Darwinism?". Proc. Natl. Acad. Sci. U.S.A. 96 (21): 11904–9.
- ↑ McGourty, Christine (2002-11-22). "Köpeklerin kökeni". BBC Haberler. 2007-12-14 (İngilizce)
- ↑ Evrimi Anlamak, Kategori sayfası: Yapay Seçilim
- 1 2 "genetik." Encyclopædia Britannica. Encyclopædia Britannica Online. Encyclopædia Britannica Inc.
- ↑ Lynch, M. (2007)."The frailty of adaptive hypotheses for the origins of organismal complexity". PNAS 104: 8597–8604.
- ↑ Smith N.G.C., Webster M.T., Ellegren, H. (2002). "Deterministic Mutation Rate Variation in the Human Genome". Genome Research 12 (9): 1350–1356.
- ↑ Petrov DA, Sangster TA, Johnston JS, Hartl DL, Shaw KL (2000). "Evidence for DNA loss as a determinant of genome size". Science 287 (5455): 1060–1062.
- ↑ Petrov DA (2002). "DNA loss and evolution of genome size in Drosophila". Genetica 115 (1): 81–91.
- ↑ Kiontke K, Barriere A , Kolotuev I, Podbilewicz B , Sommer R, Fitch DHA , Felix MA (2007). "Trends, stasis, and drift in the evolution of nematode vulva development". Current Biology 17 (22): 1925–1937.
- ↑ Braendle C, Baer CF, Felix MA (2010). "Bias and Evolution of the Mutationally Accessible Phenotypic Space in a Developmental System". PLoS Genetics 6 (3).
- ↑ Palmer, RA (2004). "Symmetry breaking and the evolution of development". Science 306 (5697): 828–833.
- ↑ West-Eberhard, M-J. (2003). Developmental plasticity and evolution. New York: Oxford University Press. ISBN 978-0-19-512235-0
- ↑ Stoltzfus, A and Yampolsky, L.Y. (2009). "Climbing Mount Probable: Mutation as a Cause of Nonrandomness in Evolution". J Hered 100 (5): 637–647.
- ↑ Yampolsky, L.Y. and Stoltzfus, A (2001). "Bias in the introduction of variation as an orienting factor in evolution". Evol Dev 3 (2): 73–83.
- ↑ Protas, Meredith; Conrad, M; Gross, JB; Tabin, C; Borowsky, R (2007). "Regressive evolution in the Mexican cave tetra, Astyanax mexicanus". Current Biology 17 (5): 452–454
- ↑ Masel J, King OD, Maughan H (2007). "The loss of adaptive plasticity during long periods of environmental stasis". American Naturalist 169 (1): 38–46.
- ↑ Lien S, Szyda J, Schechinger B, Rappold G, Arnheim N (2000). "Evidence for heterogeneity in recombination in the human pseudoautosomal region: high resolution analysis by sperm typing and radiation-hybrid mapping". Am. J. Hum. Genet. 66 (2): 557–66.
- ↑ Barton, N H (2000). "Genetic hitchhiking". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 355 (1403): 1553–1562.
- ↑ Gillespie, John H. (2001). "Is the population size of a species relevant to its evolution?". Evolution 55 (11): 2161–2169.
- 1 2 3 Scott EC, Matzke NJ (2007). "Biological design in science classrooms". Proc. Natl. Acad. Sci. U.S.A. 104 1 (suppl_1): 8669–76.
- ↑ Coyne; Barton, Turelli (1997). "Perspective: A Critique of Sewall Wright's Shifting Balance Theory of Evolution". Evolution. 3 51: 643–671.
- 1 2 3 4 5 coevolution. Berkeley Üniversitesi, erişim: 02 Kasım 2008 (İngilizce)
- ↑ Wade MJ (2007). "The co-evolutionary genetics of ecological communities". Nat. Rev. Genet. 8(3): 185–95.
- 1 2 3 Boğa boynuzlu akasya Killerplants.com. Erişim:7 Kasım 2008. (İngilizce)
- ↑ Geffeney S, Brodie ED, Ruben PC, Brodie ED (2002). "Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels". Science 297 (5585): 1336–9 *Brodie ED, Ridenhour BJ, Brodie ED (2002). "The evolutionary response of predators to dangerous prey: hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts". Evolution 56 (10): 2067–82.
- ↑ Hendry AP, Kinnison MT (2001). "An introduction to microevolution: rate, pattern, process". Genetica 112–113: 1–8.
- ↑ Leroi AM (2000). "The scale independence of evolution". Evol. Dev. 2 (2): 67–77.
- 1 2 Gould SJ (1994)."Tempo and mode in the macroevolutionary reconstruction of Darwinism". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6764–71.
- ↑ ablonski, D. (2000). "Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology". Paleobiology 26 (sp4): 15–52.
- ↑ Michael J. Dougherty. Is the human race evolving or devolving? Scientific American July 20, 1998.
- ↑ TalkOrigins Archive response to Creationist claims – Claim CB932: Evolution of degenerate forms
- ↑ Carroll SB (2001). "Chance and necessity: the evolution of morphological complexity and diversity". Nature 409 (6823): 1102–9.
- ↑ Whitman W, Coleman D, Wiebe W (1998). "Prokaryotes: the unseen majority". Proc Natl Acad Sci U S 95 (12): 6578–83.
- ↑ Schloss P, Handelsman J (2004). "Status of the microbial census". Microbiol Mol Biol Rev 68 (4): 686–91.
- ↑ Nealson K (1999). "Post-Viking microbiology: new approaches, new data, new insights". Origins of Life And Evolution of Biospheres 29 (1): 73–93.
- ↑ Buckling A, Craig Maclean R, Brockhurst MA, Colegrave N (2009). "The Beagle in a bottle".Nature 457 (7231): 824–9.
- ↑ Elena SF, Lenski RE (2003). "Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation". Nat. Rev. Genet. 4 (6): 457–69.
- ↑ Mayr, Ernst 1982. The growth of biological thought. Harvard. p483: "Adaptation... could no longer be considered a static condition, a product of a creative past and became instead a continuing dynamic process."
- ↑ The Oxford Dictionary of Science defines adaptation as "Any change in the structure or functioning of an organism that makes it better suited to its environment".
- ↑ Orr H (2005). "The genetic theory of adaptation: a brief history". Nat. Rev. Genet. 6 (2): 119–27.
- ↑ Dobzhansky, T.; Hecht, MK; Steere, WC (1968). "On some fundamental concepts of evolutionary biology". Evolutionary biology volume 2 (1st ed.). New York: Appleton-Century-Crofts. pp. 1–34.
- ↑ Dobzhansky, T. (1970). Genetics of the evolutionary process. N.Y.: Columbia. pp. 4–6, 79–82, 84–87. ISBN 0-231-02837-7.
- ↑ Dobzhansky, T. (1956). "Genetics of natural populations XXV. Genetic changes in populations of Drosophila pseudoobscura and Drosphila persimilis in some locations in California". Evolution 10 (1): 82–92.
- ↑ Nakajima A, Sugimoto Y, Yoneyama H, Nakae T (2002). "High-level fluoroquinolone resistance in Pseudomonas aeruginosa due to interplay of the MexAB-OprM efflux pump and the DNA gyrase mutation". Microbiol. Immunol. 46 (6): 391–5.
- ↑ Okada H, Negoro S, Kimura H, Nakamura S (1983). "Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers". Nature 306 (5939): 203–6.
- ↑ Ohno S (1984). "Birth of a unique enzyme from an alternative reading frame of the preexisted, internally repetitious coding sequence". Proc. Natl. Acad. Sci. U.S.A. 81 (8): 2421–5.
- ↑ Copley SD (2000). "Evolution of a metabolic pathway for degradation of a toxic xenobiotic: the patchwork approach". Trends Biochem. Sci. 25 (6): 261–5.
- ↑ Crawford RL, Jung CM, Strap JL (2007). "The recent evolution of pentachlorophenol (PCP)-4-monooxygenase (PcpB) and associated pathways for bacterial degradation of PCP". Biodegradation 18 (5): 525–39.
- ↑ Eshel I (1973). "Clone-selection and optimal rates of mutation". Journal of Applied Probability 10 (4): 728–738.
- ↑ Masel J, Bergman A, (2003). "The evolution of the evolvability properties of the yeast prion [PSI+]". Evolution 57 (7): 1498–1512.
- ↑ Lancaster AK, Bardill JP, True HL, Masel J (2010). "The Spontaneous Appearance Rate of the Yeast Prion [PSI+] and Its Implications for the Evolution of the Evolvability Properties of the [PSI+] System". Genetics 184 (2): 393–400.
- ↑ Draghi J, Wagner G (2008). "Evolution of evolvability in a developmental model". Theoretical Population Biology 62: 301–315.
- 1 2 Bejder L, Hall BK (2002). "Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss". Evol. Dev. 4 (6): 445–58.
- ↑ Young, Nathan M.; Hallgrímsson, B (2005). "Serial homology and the evolution of mammalian limb covariation structure". Evolution 59 (12): 2691–704
- ↑ Penny D, Poole A (1999). "The nature of the last universal common ancestor". Curr. Opin. Genet. Dev. 9 (6): 672–77.
- ↑ Hall, Brian K (2003). "Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution". Biological Reviews of the Cambridge Philosophical Society 78 (3): 409–433.
- ↑ Shubin, Neil; Tabin, C; Carroll, S (2009). "Deep homology and the origins of evolutionary novelty". Nature 457 (7231): 818–823.
- 1 2 3 Fong D, Kane T, Culver D (1995). "Vestigialization and Loss of Nonfunctional Characters". Ann. Rev. Ecol. Syst. 26 (4): 249–68.
- ↑ Zhang Z, Gerstein M (2004). "Large-scale analysis of pseudogenes in the human genome". Curr. Opin. Genet. Dev. 14 (4): 328–35
- ↑ Jeffery WR (2005). "Adaptive evolution of eye degeneration in the Mexican blind cavefish". J. Hered. 96 (3): 185–96.
- ↑ Maxwell EE, Larsson HC (2007). "Osteology and myology of the wing of the Emu (Dromaius novaehollandiae) and its bearing on the evolution of vestigial structures". J. Morphol. 268 (5): 423–41.
- ↑ Silvestri AR, Singh I (2003). "The unresolved problem of the third molar: would people be better off without it?". Journal of the American Dental Association (1939) 134 (4): 450–5.
- ↑ Darwin, Charles. (1872) "İnsan ve Hayvanlarda Duyguların İfadesi" John Murray, London.
- ↑ Peter Gray (2007). Psychology (fifth ed.). Worth Publishers. p. 66. ISBN 0-7167-0617-2.
- ↑ Behavior Development in Infants by Evelyn Dewey, citing a study "Reflexes and other motor activities in newborn infants: a report of 125 cases as a preliminary study of infant behavior" published in the Bull. Neurol. Inst. New York, 1932, Vol. 2, pp. 1–56.
- ↑ Jerry Coyne (2009). Why Evolution is True. Penguin Group. pp. 85–86. ISBN 978-0-670-02053.
- ↑ Anthony Stevens (1982). Archetype: A Natural History of the Self. Routledge & Kegan Paul. p. 87. ISBN 0-7100-0980-1.
- 1 2 Gould 2002, pp. 1235–6
- ↑ Piatigorsky J, Kantorow M, Gopal-Srivastava R, Tomarev SI (1994). "Recruitment of enzymes and stress proteins as lens crystallins". EXS 71: 241–50. PMID 8032155.
- ↑ Wistow G (1993). "Lens crystallins: gene recruitment and evolutionary dynamism". Trends Biochem. Sci. 18 (8): 301–6.
- ↑ Hardin G (1960). "The competitive exclusion principle". Science 131 (3409): 1292–7.
- ↑ Kocher TD (2004). "Adaptive evolution and explosive speciation: the cichlid fish model". Nat. Rev. Genet. 5 (4): 288–98.
- ↑ Johnson NA, Porter AH (2001). "Toward a new synthesis: population genetics and evolutionary developmental biology". Genetica 112–113: 45–58.
- ↑ Baguñà J, Garcia-Fernàndez J (2003)."Evo-Devo: the long and winding road". Int. J. Dev. Biol. 47 (7–8): 705–13. *Love AC. (2003). "Evolutionary Morphology, Innovation and the Synthesis of Evolutionary and Developmental Biology". Biology and Philosophy 18 (2): 309–345.
- ↑ Allin EF (1975). "Evolution of the mammalian middle ear". J. Morphol. 147 (4): 403–37.
- ↑ Harris MP, Hasso SM, Ferguson MW, Fallon JF (2006). "The development of archosaurian first-generation teeth in a chicken mutant". Curr. Biol. 16 (4): 371–7.
- ↑ Carroll SB (2008). "Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution". Cell 134 (1): 25–36.
- ↑ Paszkowski U (2006). "Mutualism and parasitism: the yin and yang of plant symbioses". Curr. Opin. Plant Biol. 9 (4): 364–70
- ↑ Paszkowski U (2006). "Mutualism and parasitism: the yin and yang of plant symbioses". Curr. Opin. Plant Biol. 9 (4): 364–70.
- ↑ Hause B, Fester T (2005). "Molecular and cell biology of arbuscular mycorrhizal symbiosis". Planta 221 (2): 184–96.
- ↑ Bertram J (2000). "The molecular biology of cancer". Mol. Aspects Med. 21 (6): 167–223.
- ↑ Reeve HK, Hölldobler B (2007). "The emergence of a superorganism through intergroup competition". Proc Natl Acad Sci U S A. 104 (23): 9736–40.
- ↑ Wilson EO, Hölldobler B (2005). "Eusociality: origin and consequences". Proc. Natl. Acad. Sci. U.S.A. 102 (38): 13367–71.
- ↑ Wilson EO, Hölldobler B (2005). "Eusociality: origin and consequences". Proc. Natl. Acad. Sci. U.S.A. 102 (38): 13367–71
- ↑ Gavrilets S (2003). "Perspective: models of speciation: what have we learned in 40 years?". Evolution 57 (10): 2197–215.
- 1 2 de Queiroz K (2005). "Ernst Mayr and the modern concept of species". Proc. Natl. Acad. Sci. U.S.A. 102 (Suppl 1): 6600–7.
- ↑ Ereshefsky, M. (1992). "Eliminative pluralism". Philosophy of Science 59 (4): 671–690.
- ↑ Mayr, E. (1942). Systematics and the Origin of Species. New York: Columbia Univ. Press.ISBN 0-231-05449-1.
- ↑ Fraser C, Alm EJ, Polz MF, Spratt BG, Hanage WP (2009). "The bacterial species challenge: making sense of genetic and ecological diversity". Science 323 (5915): 741–6.
- ↑ de Queiroz K (2005)."Ernst Mayr and the modern concept of species". Proc. Natl. Acad. Sci. U.S.A. 102 (Suppl 1): 6600–7.
- ↑ Ereshefsky, M. (1992). "Eliminative pluralism.". Philosophy of Science 59 (4): 671–690.
- ↑ Short RV (1975). "The contribution of the mule to scientific thought". J. Reprod. Fertil. Suppl. (23): 359–64.
- ↑ Gross B, Rieseberg L (2005). "The ecological genetics of homoploid hybrid speciation". J. Hered. 96 (3): 241–52.
- ↑ Vrijenhoek RC (2006). "Polyploid hybrids: multiple origins of a treefrog species". Curr. Biol. 16(7): R245.
- ↑ Rice, W.R.; Hostert (1993). "Laboratory experiments on speciation: what have we learned in 40 years". Evolution 47 (6): 1637–1653. *Jiggins CD, Bridle JR (2004). "Speciation in the apple maggot fly: a blend of vintages?". Trends Ecol. Evol. (Amst.) 19 (3): 111–4. *Boxhorn, J (1995). "Observed Instances of Speciation". TalkOrigins Archive. Retrieved 2008-12-26. *Weinberg JR, Starczak VR, Jorg, D (1992). "Evidence for Rapid Speciation Following a Founder Event in the Laboratory". Evolution 46 (4): 1214–20.
- ↑ Herrel, A.; Huyghe, K.; Vanhooydonck, B.; Backeljau, T.; Breugelmans, K.; Grbac, I.; Van Damme, R.; Irschick, D.J. (2008). "Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource". Proceedings of the National Academy of Sciences 105 (12): 4792–5.
- ↑ Losos, J.B. Warhelt, K.I. Schoener, T.W. (1997). "Adaptive differentiation following experimental island colonization in Anolis lizards". Nature 387 (6628): 70–3.
- ↑ Hoskin CJ, Higgle M, McDonald KR, Moritz C (2005). "Reinforcement drives rapid allopatric speciation". Nature 437 (7063): 1353–356
- ↑ Templeton AR (1 April 1980). "The theory of speciation via the founder principle". Genetics 94(4): 1011–38
- ↑ Gavrilets S (2003). "Perspective: models of speciation: what have we learned in 40 years?". Evolution 57 (10): 2197–215.
- ↑ Nosil P, Crespi B, Gries R, Gries G (2007). "Natural selection and divergence in mate preference during speciation". Genetica 129 (3): 309–27.
- ↑
- Savolainen V, Anstett M-C, Lexer C, Hutton I, Clarkson JJ, Norup MV, Powell MP, Springate D, Salamin N, Baker WJr (2006). "Sympatric speciation in palms on an oceanic island". Nature 441(7090): 210–3. *Barluenga M, Stölting KN, Salzburger W, Muschick M, Meyer A (2006). "Sympatric speciation in Nicaraguan crater lake cichlid fish". Nature 439 (7077): 719–23.
- ↑ Gavrilets S (2006). "The Maynard Smith model of sympatric speciation". J. Theor. Biol. 239 (2): 172–82.
- ↑ Wood TE, Takebayashi N, Barker MS, Mayrose I, Greenspoon PB, Rieseberg LH (2009). "The frequency of polyploid speciation in vascular plants". Proc. Natl. Acad. Sci. U.S.A. 106 (33): 13875–9.
- ↑ Hegarty Mf, Hiscock SJ (2008). "Genomic clues to the evolutionary success of polyploid plants".Current Biology 18 (10): 435–44.
- ↑ Jakobsson M, Hagenblad J, Tavaré S (2006). "A unique recent origin of the allotetraploid species Arabidopsis suecica: Evidence from nuclear DNA markers". Mol. Biol. Evol. 23 (6): 1217–31.
- ↑ Säll T, Jakobsson M, Lind-Halldén C, Halldén C (2003). "Chloroplast DNA indicates a single origin of the allotetraploid Arabidopsis suecica". J. Evol. Biol. 16 (5): 1019–29.
- ↑ Bomblies K, Weigel D (2007). "Arabidopsis-a model genus for speciation". Curr Opin Genet Dev17 (6): 500–4.
- ↑ Sémon M, Wolfe KH (2007). "Consequences of genome duplication". Curr Opin Genet Dev 17(6): 505–12.
- ↑ "Punctuated equilibria: an alternative to phyletic gradualism" In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82–115. Reprinted in N. Eldredge Time frames. Princeton: Princeton Univ. Press. 1985
- ↑ Benton MJ (1995). "Diversification and extinction in the history of life". Science 268 (5207): 52–8.
- ↑ Raup DM (1986). "Biological extinction in Earth history". Science 231 (4745): 1528–33.
- ↑ Avise JC, Hubbell SP, Ayala FJ. (2008). "In the light of evolution II: Biodiversity and extinction". Proc. Natl. Acad. Sci. U.S.A. 105 (Suppl 1): 11453–7.
- 1 2 3 Raup DM (1994). "The role of extinction in evolution". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6758–63.
- ↑ Novacek MJ, Cleland EE (2001). "The current biodiversity extinction event: scenarios for mitigation and recovery". Proc. Natl. Acad. Sci. U.S.A. 98 (10): 5466–70.
- ↑ Lewis OT (2006). "Climate change, species-area curves and the extinction crisis". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 361 (1465): 163–71.
- ↑ Kutschera U, Niklas K (2004). "The modern theory of biological evolution: an expanded synthesis". Naturwissenschaften 91 (6): 255–76.
- ↑ Jablonski D (2001). "Lessons from the past: evolutionary impacts of mass extinctions". Proc. Natl. Acad. Sci. U.S.A. 98 (10): 5393–8.
- ↑ Isaak, Mark (2005). "Claim CB090: Evolution without abiogenesis". TalkOrigins Archive. 14 Haziran 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150614000009/http://www.talkorigins.org/indexcc/CB/CB090.html. Erişim tarihi: 2007-05-13.
- ↑ Observed Instances of Speciation Talkorigins
- ↑ Some More Observed Speciation Events Talkorigins
- ↑ Rose, M. R. (1984). "Artificial selection on a fitness component in Drosophila melanogaster". Evolution 38 (3): 516–526. DOI:10.2307/2408701. JSTOR 2408701.
- ↑ E. coli Long-term Experimental Evolution Project Site, Lenski, R. E.
- ↑ Artificial Selection for Increased Wheel-Running Behavior in House Mice, John G. Swallow, Patrick A. Carter, and Theodore Garland, Jr, Behavior Genetics, Vol. 28, No. 3, 1998
- ↑ Peretó J (2005). "Controversies on the origin of life". Int. Microbiol. 8 (1): 23–31. PMID 15906258. http://www.im.microbios.org/0801/0801023.pdf.
- ↑ Luisi PL, Ferri F, Stano P (2006). "Approaches to semi-synthetic minimal cells: a review". Naturwissenschaften 93 (1): 1–13. DOI:10.1007/s00114-005-0056-z. PMID 16292523.
- ↑ Trevors JT, Abel DL (2004). "Chance and necessity do not explain the origin of life". Cell Biol. Int. 28 (11): 729–39. DOI:10.1016/j.cellbi.2004.06.006. PMID 15563395.Forterre P, Benachenhou-Lahfa N, Confalonieri F, Duguet M, Elie C, Labedan B (1992). "The nature of the son universal ancestor and the root of the tree of life, still open questions". BioSystems 28 (1–3): 15–32. DOI:10.1016/0303-2647(92)90004-I. PMID 1337989.
- ↑ Joyce GF (2002). "The antiquity of RNA-based evolution". Nature 418 (6894): 214–21. DOI:10.1038/418214a. PMID 12110897.
- ↑ Trevors JT, Psenner R (2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". FEMS Microbiol. Rev. 25 (5): 573–82. DOI:10.1111/j.1574-6976.2001.tb00592.x. PMID 11742692.
- ↑ Penny D, Poole A (1999). "The nature of the last universal common ancestor". Curr. Opin. Genet. Dev. 9 (6): 672–77. DOI:10.1016/S0959-437X(99)00020-9. PMID 10607605.
- ↑ Theobald, DL. (2010). "A formal test of the theory of universal common ancestry". Nature 465 (7295): 219–22. Bibcode 2010Natur.465..219T. DOI:10.1038/nature09014. PMID 20463738.
- ↑ Bapteste E, Walsh DA (2005). "Does the 'Ring of Life' ring true?". Trends Microbiol. 13 (6): 256–61. DOI:10.1016/j.tim.2005.03.012. PMID 15936656.
- ↑ Darwin, Charles (1859). On the Origin of Species (1st bas.). London: John Murray. s. 1. ISBN 0-8014-1319-2. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=text&pageseq=16.
- ↑ Doolittle WF, Bapteste E (2007). "Pattern pluralism and the Tree of Life hypothesis". Proc. Natl. Acad. Sci. U.S.A. 104 (7): 2043–9. Bibcode 2007PNAS..104.2043D. DOI:10.1073/pnas.0610699104. PMC 1892968. PMID 17261804. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1892968.
- ↑ Kunin V, Goldovsky L, Darzentas N, Ouzounis CA (2005). "The net of life: reconstructing the microbial phylogenetic network". Genome Res. 15 (7): 954–9. DOI:10.1101/gr.3666505. PMC 1172039. PMID 15965028. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1172039.
- ↑ Jablonski D (1999). "The future of the fossil record". Science 284 (5423): 2114–16. DOI:10.1126/science.284.5423.2114. PMID 10381868.
- ↑ Mason SF (1984). "Origins of biomolecular handedness". Nature 311 (5981): 19–23. Bibcode 1984Natur.311...19M. DOI:10.1038/311019a0. PMID 6472461.
- ↑ Wolf YI, Rogozin IB, Grishin NV, Koonin EV (2002). "Genome trees and the tree of life". Trends Genet. 18 (9): 472–79. DOI:10.1016/S0168-9525(02)02744-0. PMID 12175808.
- ↑ Varki A, Altheide TK (2005). "Comparing the human and chimpanzee genomes: searching for needles in a haystack". Genome Res. 15 (12): 1746–58. DOI:10.1101/gr.3737405. PMID 16339373.
- 1 2 Cavalier-Smith T (2006). "Cell evolution and Earth history: stasis and revolution". Philos Trans R Soc Lond B Biol Sci 361 (1470): 969–1006. DOI:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1578732.
- ↑ Schopf J (2006). "Fossil evidence of Archaean life". Philos Trans R Soc Lond B Biol Sci 361 (1470): 869–85. DOI:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1578735.
*Altermann W, Kazmierczak J (2003). "Archean microfossils: a reappraisal of early life on Earth". Res Microbiol 154 (9): 611–17. DOI:10.1016/j.resmic.2003.08.006. PMID 14596897. - ↑ Schopf J (1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proc Natl Acad Sci U S A 91 (15): 6735–42. Bibcode 1994PNAS...91.6735S. DOI:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=44277.
- ↑ Poole A, Penny D (2007). "Evaluating hypotheses for the origin of eukaryotes". Bioessays 29 (1): 74–84. DOI:10.1002/bies.20516. PMID 17187354.
- ↑ Dyall S, Brown M, Johnson P (2004). "Ancient invasions: from endosymbionts to organelles". Science 304 (5668): 253–57. Bibcode 2004Sci...304..253D. DOI:10.1126/science.1094884. PMID 15073369.
- ↑ Martin W (2005). "The missing link between hydrogenosomes and mitochondria". Trends Microbiol. 13 (10): 457–59. DOI:10.1016/j.tim.2005.08.005. PMID 16109488.
- ↑ Lang B, Gray M, Burger G (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annu Rev Genet 33 (1): 351–97. DOI:10.1146/annurev.genet.33.1.351. PMID 10690412.
*McFadden G (1999). "Endosymbiosis and evolution of the plant cell". Curr Opin Plant Biol 2 (6): 513–19. DOI:10.1016/S1369-5266(99)00025-4. PMID 10607659. - ↑ DeLong E, Pace N (2001). "Environmental diversity of bacteria and archaea". Syst Biol 50 (4): 470–8. DOI:10.1080/106351501750435040. PMID 12116647.
- ↑ Kaiser D (2001). "Building a multicellular organism". Annu. Rev. Genet. 35 (1): 103–23. DOI:10.1146/annurev.genet.35.102401.090145. PMID 11700279.
- ↑ Valentine JW, Jablonski D, Erwin DH (1 March 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Development 126 (5): 851–9. PMID 9927587. http://dev.biologists.org/cgi/reprint/126/5/851.
- ↑ Ohno S (1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". J. Mol. Evol.. 44 1 (S1): S23–7. DOI:10.1007/PL00000055. PMID 9071008.
*Valentine J, Jablonski D (2003). "Morphological and developmental macroevolution: a paleontological perspective". Int. J. Dev. Biol. 47 (7–8): 517–22. PMID 14756327. http://www.ijdb.ehu.es/web/paper.php?doi=14756327. - ↑ Waters ER (2003). "Molecular adaptation and the origin of land plants". Mol. Phylogenet. Evol. 29 (3): 456–63. DOI:10.1016/j.ympev.2003.07.018. PMID 14615186.
- ↑ Mayhew PJ (2007). "Why are there so many insect species? Perspectives from fossils and phylogenies". Biol Rev Camb Philos Soc 82 (3): 425–54. DOI:10.1111/j.1469-185X.2007.00018.x. PMID 17624962.
- ↑ Carroll RL (2007). "The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians". Zool J Linn Soc 150 (s1): 1–140. DOI:10.1111/j.1096-3642.2007.00246.x. PMID 12752770.
- ↑ Wible JR, Rougier GW, Novacek MJ, Asher RJ (2007). "Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary". Nature 447 (7147): 1003–1006. DOI:10.1038/nature05854. PMID 17581585.
- ↑ Witmer LM (2011). "Palaeontology: An icon knocked from its perch". Nature 475 (7357): 458–459. DOI:10.1038/475458a. PMID 21796198.
- ↑ Schloss P, Handelsman J (2004). "Status of the microbial census". Microbiol Mol Biol Rev 68 (4): 686–91. doi:10.1128/MMBR.68.4.686-691.2004. PMC 539005. PMID 15590780.
- ↑ stratigraphy.org: International Stratigraphic Chart 2009 (PDF)
- ↑ C. Brauckmann, B. Brauckmann, E. Gröning: The statigraphical position of the oldest Pterygota (Insecta. Carboniferous, Namurian). Annales de la Société géologique de Belgique 117(1), 1994, 47-56.
- ↑ G. Niedźwiedzki, P. Szrek, K. Narkiewicz, M. Narkiewicz, P. E. Ahlberg: Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463 (7277), 2010, 43–8. doi:10.1038/nature08623
- ↑ Wellman, C. H. The invasion of the land by plants: when and where? New Phytologist 188, 2010, 306-309.
- ↑ J. A. Dunlop: A trigonotarbid arachnid from the Upper Silurian of Shropshire. Palaeontology 39(3), 1996, 605-614.
P. Selden, H. Read: The oldest land animals: Silurian millepedes from Scotland. Bulletin of the British Myriapod & Isopod Group 23, 2008, 36-37. - ↑ C. V. Rubinstein, P. Gerrienne, G. S. de la Puente, R. A. Astini and P. Steemans: Early Middle Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytologist 188, 2010, 365–369. [doi: 10.1111/j.1469-8137.2010.03433.x]
- ↑ D.-G. Shu, H.-L. Luo, S. Conway Morris, X.-L. Zhang, S.-X. Hu, L. Chen, J. Han, M. Zhu, Y. Li, L.-Z. Chen: Lower Cambrian Vertebrates from South China. Nature 402(6757), 1999, 42–46. doi:10.1038/46965
- ↑ E. Landing, B.Kröger: The oldest cephalopods from East Laurentia. Journal of Paleontology, 83(1), 2009, 123–127.
- ↑ S. Nichols, G. Wörheide: Sponges: New Views of Old Animals. Integrative and Comparative Biology 45 (2), 2005, 333–334. doi:10.1093/icb/45.2.333
G. D. Love, E. Grosjean, C. Stalvies, D. A. Fike, J. P. Grotzinger, A. S. Bradley, A. E. Kelly, M. Bhatia, W. Meredith, C. E. Snape, S. A. Bowring, D. J. Condon, R. E. Summons: Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 457, 2009, 718-722 - ↑ Çağrı Mert Bakırcı. Evrim Kuramı ve Mekanizmaları Sy.34 "Kanunlar Bunun Neresinde?" Başlığı ISBN 978-605-4834-46-4
- ↑ Myers, PZ (2006-06-18). "Ann Coulter: No evidence for evolution?". Pharyngula (scienceblogs.com). http://scienceblogs.com/pharyngula/2006/06/ann_coulter_no_evidence_for_ev.php. Erişim tarihi: 2006-11-18.
- ↑ The National Science Teachers Association's position statement on the teaching of evolution.
- ↑ IAP Statement on the Teaching of Evolution Joint statement issued by the national science academies of 67 countries, including the United Kingdom's Royal Society (PDF file)
- ↑ From the American Association for the Advancement of Science, the world's largest general scientific society: 2006 Statement on the Teaching of Evolution (PDF file), AAAS Denounces Anti-Evolution Laws
- ↑ Fact, Fancy, and Myth on Human Evolution, Alan J. Almquist, John E. Cronin, Current Anthropology, Vol. 29, No. 3 (Jun., 1988), pp. 520–522
- ↑ Martz, Larry; McDaniel, Ann (1987-06-29). "Keeping God Out of Class (Washington and bureau reports)". Newsweek (Newsweek Inc.) CIX (26): 22–23
- ↑ http://www.paleogenetics.com/evodevo.html
- ↑ http://ac.els-cdn.com/S0960982206000649/1-s2.0-S0960982206000649-main.pdf?_tid=e44101ba1f5c7b708c4d43cfc27312f1&acdnat=1334307433_916a65d854a12df0e3ea650869633f59
- ↑ http://teknoloji.milliyet.com.tr/evrimin-hizi-olculdu-/uzay/haberdetay/04.01.2010/1181635/default.htm
- ↑ http://www.bilimvegelecek.com.tr/?goster=1433
- ↑ ABD Nüfus İdaresi Resmi Sitesi, James isminin görülme sıklığı verileri (İngilizce)
- ↑ ABD Nüfus İdaresi Resmi Sitesi, Steve isminin görülme sıklığı verileri (İngilizce)
- ↑ Bull JJ, Wichman HA (2001). "Applied evolution". Annu Rev Ecol Syst 32 (1): 183–217. DOI:10.1146/annurev.ecolsys.32.081501.114020.
- ↑ Doebley JF, Gaut BS, Smith BD (2006). "The molecular genetics of crop domestication". Cell 127 (7): 1309–21. DOI:10.1016/j.cell.2006.12.006. PMID 17190597.
- ↑ Jäckel C, Kast P, Hilvert D (2008). "Protein design by directed evolution". Annu Rev Biophys 37 (1): 153–73. DOI:10.1146/annurev.biophys.37.032807.125832. PMID 18573077.
- ↑ Maher B. (2009). "Evolution: Biology's next top model?". Nature 458 (7239): 695–8. DOI:10.1038/458695a. PMID 19360058.
- ↑ Borowsky R (2008). "Restoring sight in blind cavefish". Curr. Biol. 18 (1): R23–4. DOI:10.1016/j.cub.2007.11.023. PMID 18177707.
- ↑ Gross JB, Borowsky R, Tabin CJ (2009). Barsh, Gregory S.. ed. "A novel role for Mc1r in the parallel evolution of depigmentation in independent populations of the cavefish Astyanax mexicanus". PLoS Genet. 5 (1): e1000326. DOI:10.1371/journal.pgen.1000326. PMC 2603666. PMID 19119422. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2603666.
- ↑ Fraser AS (1958). "Monte Carlo analyses of genetic models". Nature 181 (4603): 208–9. Bibcode 1958Natur.181..208F. DOI:10.1038/181208a0. PMID 13504138.
- ↑ Rechenberg, Ingo (1973) (in German). Evolutionsstrategie – Optimierung technischer Systeme nach Prinzipien der biologischen Evolution (PhD thesis). Fromman-Holzboog.
- ↑ Holland, John H. (1975). Adaptation in Natural and Artificial Systems. University of Michigan Press. ISBN 0-262-58111-6.
- ↑ Koza, John R. (1992). Genetic Programming. MIT Press. ISBN 0-262-11170-5.
- ↑ Jamshidi M (2003). "Tools for intelligent control: fuzzy controllers, neural networks and genetic algorithms". Philosophical transactions. Series A, Mathematical, physical and engineering sciences 361 (1809): 1781–808. Bibcode 2003RSPTA.361.1781J. DOI:10.1098/rsta.2003.1225. PMID 12952685.
- ↑ Browne, Janet (2003). Charles Darwin: The Power of Place. London: Pimlico. ss. 376–379. ISBN 0-7126-6837-3.
- ↑ Kutschera U, Niklas K (2004). "The modern theory of biological evolution: an expanded synthesis". Naturwissenschaften 91 (6): 255–76. Bibcode 2004NW.....91..255K. doi: 10.1007/s00114-004-0515-y. PMID 15241603.
- ↑ For an overview of the philosophical, religious and cosmological controversies, see: Dennett, D (1995). Darwin's Dangerous Idea: Evolution and the Meanings of Life. Simon & Schuster. ISBN 978-0-684-82471-0.
*For the scientific and social reception of evolution in the 19th and early 20th centuries, see: Johnston, Ian C.. "History of Science: Origins of Evolutionary Theory". And Still We Evolve (Liberal Studies Department, Malaspina University College). http://records.viu.ca/~johnstoi/darwin/sect3.htm. Erişim tarihi: 2007-05-24.
*Bowler, PJ (2003). Evolution: The History of an Idea, Third Edition, Completely Revised and Expanded. University of California Press. ISBN 978-0-520-23693-6.
*Zuckerkandl E (2006). "Intelligent design and biological complexity". Gene 385: 2–18. DOI:10.1016/j.gene.2006.03.025. PMID 17011142. - ↑ Scott EC, Matzke NJ (2007). "Biological design in science classrooms". Proc. Natl. Acad. Sci. U.S.A.. 104 1 (suppl_1): 8669–76
- ↑ Ross, M.R. (2005). "Who Believes What? Clearing up Confusion over Intelligent Design and Young-Earth Creationism". Journal of Geoscience Education 53 (3): 319. http://www.nagt.org/files/nagt/jge/abstracts/Ross_v53n3p319.pdf. Erişim tarihi: 2008-04-28.
- ↑ Hameed, Salman (2008-12-12). "Science and Religion: Bracing for Islamic Creationism". Science 322 (5908): 1637–1638. DOI:10.1126/science.1163672. PMID 19074331. http://helios.hampshire.edu/~sahCS/Hameed-Science-Creationism.pdf. Erişim tarihi: 2009.
- ↑ Bowler, Peter J. (2003). Evolution:The History of an Idea. University of California Press. ISBN 0-520-23693-9.
- ↑ Spergel D. N.; Scott, EC; Okamoto, S (2006). "Science communication. Public acceptance of evolution". Science 313 (5788): 765–66. DOI:10.1126/science.1126746. PMID 16902112.
- ↑ Understanding Creationism after Kitzmiller 2007
- ↑ Religious Differences on the Question of Evolution, Pew Forum, 2009-2-4
- 1 2 nationalgeographic.com
- ↑ Science 11. Ağustos. 2006 tarihli 5788. sayısı, cilt 313
- 1 2 Dr. Kenan Ateş, İkili Sarmal - Evrim neden ABD ve Türkiye'de az benimseniyor?, Bilim ve Gelecek, Dergi arşivi, Sayı 31
- ↑ Hakan Çetinkaya, "Evrim, Bilim ve Eğitim Üzerine", Eğitim Fakültesi, Ege Eğitim Dergisi, 2006 (7) 1:1-21
- ↑ Okt. Osman SAÇIKARA, Safsata Anketi ve Bilimsel Düşünce, Adnan Menderes Üniversitesi, 16 Aralık 2004
- ↑ 59% dos brasileiros acreditam em Deus e também em Darwin
- ↑ Angus Reid Polls
- ↑ Britons unconvinced on evolution
- ↑ BBC Survey On The Origins Of Life
- ↑ JACQUELINE MALEY (2009-12-19). God is still tops but angels rate well. The Age. http://www.theage.com.au/national/god-is-still-tops-but-angels-rate-well-20091218-l5v9.html
- ↑ Science literacy in Australia Federation of Australian Scientific and Technological Societies and Australian Academy of Science (PDF file)
Dış bağlantılar
- Evrimi Anlamak Berkeley Üniversitesinin "Understanding Evolution" isimli sitesinin Türkçe çevirisi
- Darwin´in bütün eserleri (İngilizce)
- Evrim Çalışkanları Evrim üzerine arşiv yapan bir blog
- Tiktaalik'in babası da bulundu
- Time Dergisi'nin "What Makes Us Different" adlı makalesinin Türkçe tercümesi (Bizi Farklı Kılan Nedir?)
- Evrim Teorisi'nin Türkiye'deki bilinci hakkında bir eleştiri
- Eğitim Fakültesi Dergisi - Biyoloji öğretim programında evrim kuramı
Videolar
- Dünyanın oluşumu ve yaşamın başlangıcı - Çİzgifilm
- Evrim gerçeği
- Balinaların evrimi
- "1 dakikada Evrim"
- Evrim teorisi karşıtı 10 iddiaya cevaplar
- Bir evrim simülasyonu
Basından ilgili haberler
- Radikal: Evrimde kayıp halka bulundu Hürriyet(2)
- Hürriyet: Evrimin moleküler temelde kanıtlanması
- Hürriyet: İnsanın köklerinin tarihi yeniden oluşuyor
- Hürriyet: Sudan karaya geçişte en önemli kayıp halka bulundu: Gogonasus
- National Geographic: Was Darwin Wrong?
Kitaplar
- Türlerin Kökeni - Charles Darwin, Janet Browne (Versus Kitap, Ağustos 2008)
- Yaşamın Tüm Çeşitliliği, İlerleme Mitosu - Stephen Jay Gould (Versus Kitap, Ocak 2009)
- Charles Darwin'in Özyaşam Öyküsü - Derleyen Francis Darwin (Daktylos Yayınları, Nisan 2009)
- Kör Saatçi - Richard Dawkins, (Tübitak Yayınları)
- Gerçeğin Büyüsü - Richard Dawkins (Editör: B. Duygu Özpolat) (Kuzey Yayınları, Kasım 2012)
- Gen Bencildir - Richard Dawkins (Kuzey Yayınları)
- Evrim Atlası - Peter Barrett, Douglas Palmer (İş Bankası Kültür Yayınları)
| |||||||||||||||||||||||||||||||||||||